
![Populasyon Genetiği]() Yeni Alellerin Ortaya Çıkması
Yeni Alellerin Ortaya Çıkması
 Yeni Alellerin Ortaya Çıkması
Yeni Alellerin Ortaya ÇıkmasıDarwin ve ilk taraftarları için, evrimin sorunlu sırlarından birisi, var olan özelliklerin küçük varyantları üzerinden kompleks bir organ ya da bir yapının seçilimle nasıl ortaya çıkabileceğidir. Örneğin, omurgalıların oldukça ayrıntılı kamera gözü nasıl evrimleşebildi? Tüm ara basamaklar ve aynı şekilde ilk adım kullanışsız ve hatta zararlı mı olur?
Önceden var olan bazı yapıların değişmesinden oluşacak her şeyin her zaman değerli olmayacağının teminatı var mıdır? Bu husus, moleküler düzeyde bir proteindeki aminoasit değişim dizisi olarak varsayılabilir mi? Bu durumda, örneğin değişimler var olan bir enzimin özgüllüğünü ve aktivitesini bozacak mı? Çok önceden, mutasyonların yeni ve daha iyi bazı kombinasyonları üretme şansı olmaz mı ? Bugün biliyoruz ki, Darwin’in genel düşüncesi doğruydu. Yeni genler, yeni aleller ve yeni morfolojik yapılar, genellikle daha önce var olanlardan bu gün iyi bilinen kalıtsal mekanizmalarla ortaya çıkarlar.
Mutasyonlarla ilgili bugünkü bilgilerimiz -baz yitirilmesi, bazların yer değiştirmesi ve bir ya da birkaç bazın eklenmesi- bu seviyedeki değişimlerin iki yararlı etkisinin olabileceğini gösterir. Birincisi, diployitliğin oluşturduğu hassas koruma ile ara kademelerde negatif seçilimden korunan, daha önceden var olan bir genin bir alelini daha iyi bir niteliğe dönüştürebilir (genin sabit versiyonu kromozomun diğer kopyasındadır ve işlevini görür). İkincisi, eğer nokta mutasyonlar kontrol bölgelerinde oluşursa (örneğin transkripsiyon faktörlerinin bağlanacağı bölgeler), bu mutasyonlar kromozomun hangi kolunda bulunuyorsa o kol üzerinde bulunan bir ya da bir grup alelin özgüllüğünü, zamanlamasını ve genin aktivitesinin derecesini tam olarak değiştirebilir. Bunların varyasyonun yararlı kaynakları olduğu kesindir, ancak daha büyük evrimsel değişimlerin kökeninde başka iki mekanizma yatmaktadır. Bu iki mekanizma (a) genin yedek kopyasının bağımsız evrimleşmesini izleyen gen duplikasyonu ve (b) önceden var olan eksonların yeniden düzenlenmesidir.
Gen ve Ekson Duplikasyonu![Populasyon Genetiği]()
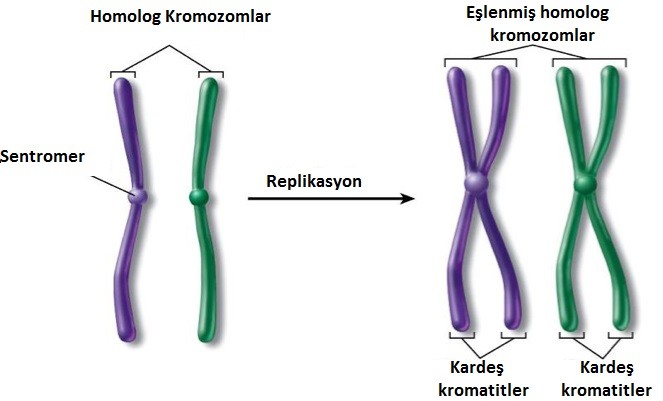
Ökaryotik kromozomların intronlar, eksonlar, görünüşte fonksiyonsuz yalancıgenler ve tekrarlı DNA dizilerinden oluştuğunun ortaya çıkarılması büyük bir sürpriz olmuştur. Biyologların çoğu, prokaryotlarda olduğu sanılan durum gibi, doğal seçilimin görünüşte gereksiz olan DNA’yı temizlemesi yönünde işlemesini beklemektedirler. Örneğin, E. coli bakterisi, doğal habitatında her zaman bol bulunan bir kaç amino asidi sentezleme yeteneğini yitirmiştir. Bu bakteride biyosentetik yollarla ilişkili genler kayıp olmuştur. Özünde, hızlı bir şekilde çoğalan bu organizmanın kromozumunda kullanılmayan bir bölge yoktur. Bunun aksine, ökaryotik DNA’nın % 90’dan daha fazlası görünüşte fonksiyonsuz ve dormant haldedir. Sonraki çalışmalarla ortaya konan, fetal immün sisteminin gereksiz gen segmentlerini aktif olarak çıkarması beklenmemekteydi ve bu da genomun düşünülmüş olduğu kadar sabit olmasının gerekmediğini ortaya koymuştur. Transpozonların keşfi bu noktanın önemini kavratmıştır.
MHC proteinleri ve T- hücre reseptörleri ve antikor genlerinin kendilerine özgü yapı ve organizasyonu genelde genlerin evrimi konusundaki düşüncelere kaynak sağlamaktadır. Yaygın olarak kabul edilen görüşlerden biri, hafif ve ağır antikorların hem ağır hem de hafif zincirlerinin değişken ve sabit bölgelerini kodlayan (bir birine çok benzeyen polipeptidler kodlarlar) eksonların önceki gen dizilerinin (ilkin gen dizilerinin) -belki hücre adhezyon moleküllerinden birinin- duplikasyonu ve düzenlenmesi ile ortaya çıktığı şeklindedir. Hücre adhezyon moleküllerinin kendilerinin çok daha eski bir genden duplikasyonla türediğine inanılan yakın bir gen familyasının bir bölümü olduğunu biliyoruz. Eksonların (ve bunlara yakın olarak intronların) ya da tam bir genin duplikasyonu, çok iyi bilinen birçok yolla oluşabilir. Bunların çoğu kromozomal anomaliliklerini gerektirir. Biri kazara oluşan mRNA geri transkripsiyonunu izleyen bir kromozoma cDNA girişi ile sonuçlanan inkorporasyondur; diğeri transpozonlarca oluşturulan duplikasyondur.
 Gen ya da eksonların duplikasyonunu izleyen küçük evrimsel değişimlerle ekstra kopyaların, yeni özellikleri verecek fonksiyonel genlerin oluşumuna yol açması olasılığının, tek başına olabilecek rasgele değişimlerle olma olası lığından daha fazla olacağını anlamak oldukça kolaydır. Harflerin ve aralıkların rasgele düzenlenmesi ile anlamlı bir cümle yaratabilme olasılığının ne kadar küçük olduğunu düşünün. Halbuki, kelimeleri ve aralıkları bulunan anlamlı bir cümle ile başlarsak (fonksiyonel bir protein için bir gen) ve var olan harflerin birkaçını değiştirirsek, bu küçük değişimlerle yeni ve oldukça anlamlı olabilecek ilginç bir cümle oluşturabiliriz.
Gen ya da eksonların duplikasyonunu izleyen küçük evrimsel değişimlerle ekstra kopyaların, yeni özellikleri verecek fonksiyonel genlerin oluşumuna yol açması olasılığının, tek başına olabilecek rasgele değişimlerle olma olası lığından daha fazla olacağını anlamak oldukça kolaydır. Harflerin ve aralıkların rasgele düzenlenmesi ile anlamlı bir cümle yaratabilme olasılığının ne kadar küçük olduğunu düşünün. Halbuki, kelimeleri ve aralıkları bulunan anlamlı bir cümle ile başlarsak (fonksiyonel bir protein için bir gen) ve var olan harflerin birkaçını değiştirirsek, bu küçük değişimlerle yeni ve oldukça anlamlı olabilecek ilginç bir cümle oluşturabiliriz.
Antikor genlerinde gördüğümüze benzer eksonların fazla sayıda çoklu kopyasının olası kaynaklarından biri duplikasyondur. İntronlar eksonların yeni kopyalarının insersiyonu (araya girmesi) için en iyi yerlerdir. Örneğin, eğer bir ekson diğer eksonların arasına girerse, belirli amino asitleri kodlayan kodonlarda, bu insersiyondan dolayı şans eseri yeni bir dizilim oluşabilir ve sonraki kodonların tümü sırasıyla kayacak ve translasyon sırasında yanlış okunacaktır. Eğer bir ekson bir intronun içine girerse, bu sorun ortaya çıkmayacaktır. mRNA oluşumu sırasında, intronlar, başlangıç ve bitiş uçlarına bağlanmış intron parçalarına bağlı snRNP’nin yardımı ile uzaklaştırılır. İntronların arasına başarı ile girmiş olan yeni eksonların transkript sürecine şifre oluşturmak üzere, koruyucu intronlarla birlikte alınması olasıdır.
Antikor genlerinde görülen bu organizasyon sonucu, değişik eksonlar, aynı özgün dizilimden köken almış olsalar bile, mutasyon ve seçilimden bağımsız olarak değişikliğe uğrayabilirler. Doğal olarak, bu değişken bölgenin birbirinden çok az farklılık gösteren, çok sayıda alternatif eksonları yaratmada da oldukça önemlidir.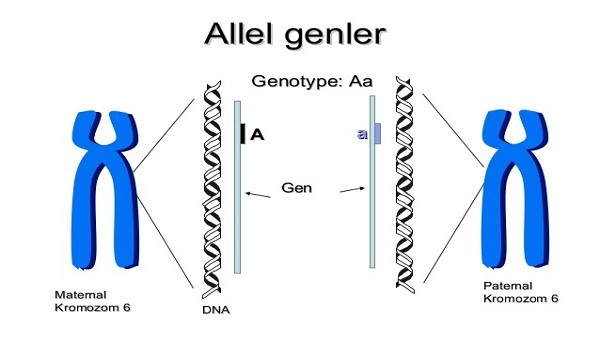
Antibadi olmayan proteinleri kodlayan genlerin duplikasyonu için kanıtlar
Omurgalıların bağışıklık sistemlerinin, çeşitli antikor üretme yeteneği, ekson duplikasyonunun daha önce geçirmiş olduğu evrimsel yolda karşı karşıya kaldığı saldırılara (vücut savunması açısından) bağlıdır. Fakat, duplikasyon kısmen de olsa her yerde enzim ve yapısal proteinleri kodlayan genlerin evriminden sorumlumudur? Miyoglobin ve hemoglobin genleri için duplikasyonun rolü ile ilgili kanıtlar hemen hemen tamdır. Kasın oksijen depo pigmenti olan miyoglobin bir polipeptid zincirinden oluşmasına karşın, kanda oksijen taşıyan protein olan Hemoglobin, iki çift, yani toplam 4 zincir içerir. Hemoglobindeki a ve b zincirlerinin üç boyutlu konformasyonu hemen hemen aynıdır, ve yine bu zincirler miyoglobindeki tek zincirin konformasyonuna çok benzerler. Bu zincirlerin tümünün genlerinin tek bir atasal genden duplikasyonla evrimleştiği düşünülmektedir. Bu görüş, bu genlerin tümünde intronların aynı bölgelerde yerleşmiş olduklarının keşfi ile güçlendirilmiştir.
Hemoglobinin kendisi, bir organizmanın yaşamında farklı dönemlerde (örneğin embriyonik gelişim dönemde ve ergin dönemde) çok az farklılık gösteren formlarda sentezlenir ve böylece farklı pH ve oksijen konsantrasyon koşulları için özelleşir. Yine, insanlarda hemoglobinin bu alternatif formlarını kodlayan genler 7. Kromozomda yan yana bulunurlar. Bu genlerin duplikasyonla oluştuğu, sonra bağımsız olarak evrimleştiği düşünülmektedir. Çünkü, aynı bölge, hemoglobin genlerininkine çok benzeyen dizilimlere sahip çok sayıda yalancı gen içermektedirler. Belki bu yalancı genler, evrimin fonksiyonel gen ürünleri düzenlenmesine yol açmadığı sonraki duplikasyonlardır.
Aynı modeller çok sayıda enzim grubunda da görünmektedir. Örneğin, sindirim enzimleri tripsin, kimotripsin ve elestaz ve kan pıhtılaşma enzimi trombin gibi enzimlerin tümü farklı fonksiyonlara sahiptir. Ancak, bu enzim genlerinin baz dizisi ve intron yerleşimi hemen hemen aynıdır. Bu genlerin her birinin, bağımsız olarak, diğerlerinin bir tekrarına yakın bir duplikantı şeklinde evrimleşmesi hemen hemen imkansızdır. Çoğu araştırıcılar, bu enzimlerden sorumlu genlerinin bir primordial (atasal) enzimi kodlayan bir geninin duplikasyonları ile herbirinin ayrı ayrı olarak farklılaştığı ve daha sonra ayrı olarak bağımsız evrimsel bir yol izlediklerine inanılmaktadır.
Okaryotik gen evriminin kısmen gen duplikasyonuna bağlı olduğu, bu gen duplikasyonunu takiben fonksiyonel olarak farklı ürünler meydana getiren baz değişimlerinin oluştuğuna ilişkin önemli kanıtlar vardır. Fakat, böylesi değişimlerin yavaş olduğu ve çoğu rasgele mutasyonun, değişmiş özgüllükten ziyade, genlerde, indirgenmiş fonksiyonlu ya da tamamen fonksiyonsuz ürünlerle sonuçlandığı unutulmamalıdır. işlevsel bir protein üreten her yeni gen için duplikasyonları içeren, tamamlanmamış ya da başarısız olmuş onlarca ya da yüzlerce teşebüs olabilir.
 Kısaca, yararsız duplikasyonları düzeltici bazı mekanizmalar işlev görmediği sürece, bir ökaryotik kromozomun, işlevsel geninkine çok benzeyen, fakat kendisi işlevsiz olan baz dizimleri ile dolu olması beklenirdi. Gerçekten gördüğümüz gibi memeli genomunun % 90’nından fazlası fonksiyonel ürünler kodlamaz. Okaryotik kromozomlann çok fazla miktardaki fonksiyonsuz yalancı genlerin, asla fonksiyonel genlere evrimleşmeyen sonraki duplikasyonların kanıtları olabilir. Tekrarlı DNA’nın birçok örneği bu kategoriye uygundur.
Kısaca, yararsız duplikasyonları düzeltici bazı mekanizmalar işlev görmediği sürece, bir ökaryotik kromozomun, işlevsel geninkine çok benzeyen, fakat kendisi işlevsiz olan baz dizimleri ile dolu olması beklenirdi. Gerçekten gördüğümüz gibi memeli genomunun % 90’nından fazlası fonksiyonel ürünler kodlamaz. Okaryotik kromozomlann çok fazla miktardaki fonksiyonsuz yalancı genlerin, asla fonksiyonel genlere evrimleşmeyen sonraki duplikasyonların kanıtları olabilir. Tekrarlı DNA’nın birçok örneği bu kategoriye uygundur.
Ekson Rekombinasyonu
Bir genin duplike olabileceğini ve sonra evrimleşerek fonksiyonel olarak farklı bir ürünü kodlayabileceğini biliyoruz. Gen dizilerinin tekrarlı duplikasyonları ile sonucu ortaya çıkmış gibi görünen antikor genlerinin rasgele seçilimini ve çeşitli antikorlar üretmek üzere kombine olan eksonlar içerdiği de aşikar. Bu gözlemler, Harward’dan Walter Gilbert ve Oxford’dan Colin Blake’in yeni genlerin nasıl evrimleşebileceği diğer bir yoluna destek sağlar. Olasılıkla farklı genlerden gelen farklı eksonlar yeni kombinasyonlar oluşturmak üzere birlikte taşınabilir.
Eğer eksonlar “domainler” (değişik şekilleri oluşturan bloklara benzeyen farklı alt üniteler, yeni bir tarzda yerleştirildiklerinde yeni yapılar oluşturan) olarak adlandırılan bir kısım sonuç proteinleri kodluyorlarsa bu yaygın olarak uğraşılan varsayım anlamlı olacaktır.
Düzinelerce proteinin genleri ile yapılan dikkatli denemeler bunun sıklıkla olan durum olduğunu göstermiştir. Örneğin, miyogloblini kodlayan gendeki her iki intron, bu çok fazla kıvrılmış globüler proteinin ana kıvrımlarda, bağlanan kısımları kodlayan bölgeleri arasında bulunur. Böylece her intron tam domainler arasında bulunan bir tür sınırı tanımlar ve her bir miyoglobin eksonunun, bu domainlerden ya da alt ünitelerden birini kodladığı düşünülebilir. Diğer bazı proteinlerin genlerinde, intronlar, x-heliks kısmını kodlayan bölgeler ile b-kıvrım tabakası kısmını kodlayan bölgelerin sınırlarında bulunur. Alternetif olarak, intronlar bazen proteinin aktif kısmını içeren kodon bölgelerinin yanında bulunur. Eksonlarca kodlanan bölgeler, farklı üniteler oluşturduklarından, eksonların yeni bir rekombinasyonu, yeni özellikleri olan çalışan bir enzimi oluşturma şansına sahip olacaktır. Sözgelimi, A geninin 2. eksonu, B geninin 4. eksonu, C geninin 1. ve 2. eksonu ve D geninin 2. eksonunu içeren yeni bir gen oluşabilir. Böyle bir rekombinasyon, eksonların orijinal genlerinden ya da duplikatlarından ayrılmasından etkilenebilir. Bu durum, eksonların kendileri ya da kopyalarının ayrılmasını da içerebilir. Her durumda kromozomun intron bölgesinin içine ekson (bağlantılı intronu ile birlikte) girişi fonksiyonel bir yeni oluşum şansını arttırır.
Son yıllarda yapılan karşılaştırmalı DNA dizin çalışmaları, geçmişteki olası ekson rekombinasyonları için çok daha fazla kanıt sağlamıştır. Gilbert, bugün var olan genlerin tümünün (yalnız insanda 50.000, binlerce eksonun yüzlerce bileşimi) büyük bir olasılıkla 1000 kadar farklı eksondan evrimleştiğini tahmin etmiştir. Daha çarpıcı olan, yeni enzimler oluşturmak için moleküler biyologlarca geliştirilen bu evrimsel senaryonun, hibrit düzenleyici genleri oluşturmak için ekson rekombinasyonunda kullanılmasıdır. Ekson duplikasyon ve rekombinasyon süreçlerinin ana evrimsel yenilikler oluşturan etkin mekanizmalar olduğu hemen hemen kesin görünmektedir.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç