
![Yaşamın Kökeni: Polimerler ve İlkel Hücrelerin Oluşması]() Polimerlerin Oluşması
Polimerlerin Oluşması
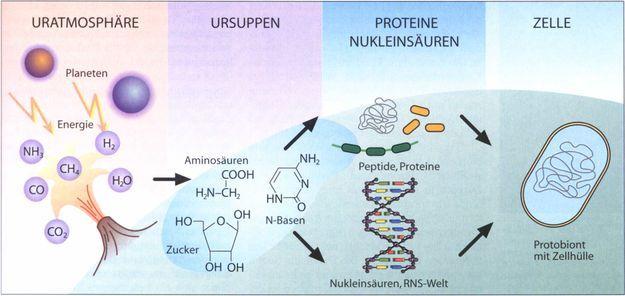
İlkin denizlerde, çeşitli hidrokarbonlar, yağ asitleri, amino asitler, pürin ve primidin bazları, basit şekerler ve nisbeten küçük diğer organik moleküllerin yavaş yavaş biriktiğini varsayalım. Bu durum yaşamın başlangıcı için yine de yeterli değildir. Özellikle polipeptitler ve nükleik asitler başta olmak üzere makromoleküllere gereksinim vardır. İlkin okyanuslarda bu polimerler “bu çorbada” bulunan yapıtaşlarından nasıl oluşturabilmişlerdir. Bu soruyu yanıtlamak kolay değildir ve günümüzde farklı araştırmacılar tarafından savunulan birçok hipotez vardır. Bazıları, denizlerdeki organik madde derişiminin, yüz milyonlarca yıllık bir periyot sonucunda, önemli miktarlarda makromolekül oluşturmak üzere, basit moleküllerin şans eseri bağlanmasına yetecek miktarda olduğunu düşünmektedir. Protein enzimler olmadan, böylesi polimerizasyon tepkimelerinin olması mümkün görülmezse bile, bu görüşün savunucuları, yine de, bu zaman skalasında, nadir ve olasılığı düşük olayların birlikte olabileceğine işaret etmektedirler. Ancak, diğer araştırmacılar, ilkin okyanuslarda organik maddenin rasgele polimerizasyon şansı için yeterince bol olduğu fikrine katılmakta isteksizdirler. Bu araştırmacılar, kimyasal tepkimelerin konsantrasyon mekanizmalarına bağlı olarak hızlandığını öne sürmüşlerdir. Böyle bir mekanizma, yapıtaşı (monomerlerin) kil parçacıkları gibi parçacıkların yüzeylerince absorbe edilmeleri olabilir. Diğer bir mekanizma, küçük miktarlardaki yapı taşı bileşiklerin çok seyreltik çözeltilerinin lagün ve gölcük sahillerindeki küçük su çukurluklarında birikimi olabilir. Güneş ısısı, suyun çoğunu buharlaştırarak organik kimyasalları yoğunlaştırmış ve hem de polimerizasyon tepkimeleri için enerji sağlamıştır. Oluşan polimerler, sonra tekrar gölcüklere geçmiş olmalıdır. Böyle bir süreç, gölcüklerde yavaş bir şekilde bir makromolekül kaynağı oluşturabilir. Sidney W. Fox tarafından gösterildiği gibi, hemen hemen kuru bir amino asit karışımı ısıtıldığında, özellikle fosfat varlığında, hızlı bir şekilde polipeptit molekülleri sentezlenebildiğinden, bu hipotez mantıklı bir hipotez olarak görülmektedir. Alternatif olarak, buharlaşma ile yoğunlaştıktan sonra gölcüklerdeki polimerizasyon tepkimeleri için gerekli enerji, ısıdan ziyade UV radyasyonundan gelmiş olabilir.
Periyodik polimerizasyon tepkimelerinde, birden fazla yoğunlaştırıcı mekanizma birlikte rol almış olsa da, daha zor anlaşılabilecek şey, yapı taşı moleküllerinin polimerler şeklinde birleşmesidir. En azından aminoasitlerin proteinler şeklinde polimerleşmesi durumu buna örnektir.
Miller ve Fox’un yapmış olduklarına benzer deneylerde, değişik araştırmacıların, ilk olarak polimerleri daha sonra da hidrolizle onların monomerlerini bulmuş olmaları şaşırtıcıdır.
Moleküler Yığılmalar ve İlkel Hücrelerin Oluşumu![Yaşamın Kökeni: Polimerler ve İlkel Hücrelerin Oluşması]()

Şimdi, yaşamın kökeni ile ilgili modelimizde, ilkin denizlerde ya da en azından nehir ağzı ve lagünlerde, tuzlar, polipeptit ve belki nükleik asit gibi polimerleri içeren organik molekül karışımından oluşmuş bir “çorbanın” olduğu bir noktaya ulaştık. Şimdi sorulması gereken şey, bu karışımdan canlı varlıklar’ karakterize edecek düzenlemenin nasıl ortaya çıktığıdır.
Oparin, uygun sıcaklık, iyonik kompozisyon ve pH koşullarında, makromolekül kolloyitlerinin (stabil süspansiyonlar) koaservat damlacıkları denen karmaşık birimleri verme eğiliminde olduğuna işaret etmiştir. Böylesi her bir damlacık, bir su kabuğu ile sınırları belirlenmiş, büyük oranda hidrofobik bir makromolekül kümesidir. Dolayısıyla, koaservat damlacığı ile damlacığın içinde yüzdüğü sıvı arasında, bir sınır ve ara yüzey vardır. Bir anlamda yüzeydeki su moleküllerinin oluşturduğu kabuk, damlacık etrafında bir membran oluşturur. Artık, koaservat damlacıkları, kendisini çevreleyen çözeltiden, hidrofilik olan değişik maddeleri tamamen ya da kısmen absorbe etme ve bünyelerine katma yönünde belirgin bir eğilime sahiptirler. Bazen bu seçici eğilim o kadar belirgindir ki, damlacıklar, ortamdaki bazı maddelerin tümünü alırlar. Bu yolla koaservat damlacığı, etrafındaki sıvının zararına büyür. Yine hidrofilik ve iyonik etkileşimlerin bir sonucu olarak, koaservat damlacıkları, belirli bir içsel yapı oluşturma yönünde güçlü bir eğilime sahiptir. Bu, damla içindeki moleküllerin rasgele dağılmış olması yerine düzenli bir tarzda yığılma eğiliminde olması demektir. Damlacık içine giren madde miktarı arttıkça, dizili su molekülleri, bu kabuğunun hemen iç yüzeyinde, yüzey aktif madde (fosfolipidlere analog) içeren bir membran oluşturabilir.
Bu şekilde damlacık sınırının geçirgenliği, öncekinden daha da seçici olmaya başlar. Böylece, koaservat damlacıkları, kelimenin tam anlamıyla canlı olmasalar bile, iç kimyasal yapılarını kendi çevrelerinden koruduklarından, canlı organizmalara ilişkin birçok özelliği gösterirler. Gerçekten, ışık mikroskobu ya da hatta elektron mikroskobu ile bakıldığında, bu tip damlacıklar, organizmalara o kadar benzerdirler ki, bazen deneyimli biyologlar bile yanlışlıkla bakterilere benzetir ve tür olarak tanımlamaya kalkışırlar.
Fox, Oparin gibi, ilk hücrelerin gelişimini sağlayan prebiyolojik sistemleri mikroskobik multimoleküler damlacıklar olarak tasarlamıştır. Ancak, Fox, koaservat damlacıklarından ziyade protenoyit mikrosferler (protenoid microspheres) olarak önermiştir. Fox’un mikrosferleri, sıcak polipeptit solüsyonları soğutulduğunda, kendiliğinden oluşan damlacıklardır. Mikrosferlerin, hipotonik bir ortamda şişmesi ve hipertonik bir ortamda büzüşmesi, içsel hareket yapabilmesi, büyüme ve karmaşıklığını artırabilmesi, yüzeysel olarak bir anlamda mayalarda görülen üreme tarzında tomurcuklanmayı andıran boğumlar meydana getirmesi ve birçok bakteride görüldüğü gibi değişik tarzlarda gruplanma şeklinde bir yığılma göstermeleri, bir hücreye özgü özellikler olduğu söylenebilir. Gerçekten, son zamanlarda, Fox, bu mikrosferleri güneş ışığına tuttuğunda, mikrosferin membranlarına karşı, hemen tüm canlı hücrelerde görülenden farklı olmayan, elektriksel bir potansiyel geliştiğini bulmuştur.
 Her iki damlacık -karmaşık koaservat ya da protenoyit mikrosfer yapısal olarak organize olduklarından ve dış ortamdan kesin olarak ayrıldıklarından, damlacığın içinde meydana gelen kimyasal tepkimeler, yalnızca ortamın koşullarına bağlı olmayacak, aynı zamanda damlacığın kendi içindeki organizasyonuna da bağlı olacaktır. Değişik maddeler, damlacık içinde daha yoğun olarak yer alabileceğinden dolayı, bu maddelerin kimyasal tepkimelerde yer alma olasılığı artar ve damlacık içindeki organizasyondan dolayı, meydana gelen her tepkime, maddeler dış ortamda serbest olarak bulunduklarından çok daha farklı şekillerde bulunacaklarından, bir ölçüde diğer tepkimeleri de etkileyeceklerdir. Ayrıca, hem metalik bileşikler gibi inorganik maddelerin hem de protein gibi organik bileşiklerin katalitik aktivitesi, moleküllerin damlacık içindeki düzenli uzaysal dizilimleri ile arttırılır. Kısaca, damlacık içindeki özel koşullar, damlacık içinde meydana gelen tepkimeler üzerinde seçici ve düzenleyici bir etki gösterecektir.
Her iki damlacık -karmaşık koaservat ya da protenoyit mikrosfer yapısal olarak organize olduklarından ve dış ortamdan kesin olarak ayrıldıklarından, damlacığın içinde meydana gelen kimyasal tepkimeler, yalnızca ortamın koşullarına bağlı olmayacak, aynı zamanda damlacığın kendi içindeki organizasyonuna da bağlı olacaktır. Değişik maddeler, damlacık içinde daha yoğun olarak yer alabileceğinden dolayı, bu maddelerin kimyasal tepkimelerde yer alma olasılığı artar ve damlacık içindeki organizasyondan dolayı, meydana gelen her tepkime, maddeler dış ortamda serbest olarak bulunduklarından çok daha farklı şekillerde bulunacaklarından, bir ölçüde diğer tepkimeleri de etkileyeceklerdir. Ayrıca, hem metalik bileşikler gibi inorganik maddelerin hem de protein gibi organik bileşiklerin katalitik aktivitesi, moleküllerin damlacık içindeki düzenli uzaysal dizilimleri ile arttırılır. Kısaca, damlacık içindeki özel koşullar, damlacık içinde meydana gelen tepkimeler üzerinde seçici ve düzenleyici bir etki gösterecektir.
İlkin dünyanın denizlerinde, oldukça fazla sayıda bu tip farklı prebiyolojik sistem oluşmuş olabilir. Büyük bir olasılıkla bunların çoğu uzun süre yaşayamayacak kadar kararsız olmuş olsa bile, bazı protenoyit mikrosferler laboratuvar koşullarında altı yıldan daha fazla bir süre kararlı kalmıştır. Bu süre, kimyasal tepkimelerin oluşması, birçok “döllük” büyümenin ve tomurcuklanmanın olması ve doğal seçilimin işlemesi için oldukça uzun bir zamandır. Bazı damlacıkların tercih edilir belirli madde kombinasyonları, özellikle katalitik aktivitesi olan kompleksler içeriyor olmaları damlacık içinde oluşan tepkimeler arasında, beklenmedik harmonik etkileşimleri geliştirmiş olabilir. Böylesi damlacıkların büyüklükleri artacağından, özellikle, daha sonra asıl damlacığın kompozisyonuna ve özelliklerine sahip, daha küçük damlacıklar verecek fiziksel bölünmeler için daha başarılı olacaklardır. Bu damlacıklar, tekrar tekrar büyümüş ve bölünmüşlerdir.
Bileşenleri abiyotik koşullarda sentezlenebilse ve bazı prebiyontların içine girebilse de, bu ilkel üreme, başlangıçta nükleik asitlerin denetiminde gerçekleşmemiş olabilir. Bu senaryodan sonraki temel adım, organik katalizörlerin işe karışması ve bunların gelecek “döner” için kopya edilme yeteneklerinin gelişme evresidir. İntronları uzaklaştıran RNA temelli enzimlerin -ribozimlerin- bulunuşu, köken olarak RNA’nın hem enzim hem de genetik kütüphane işlemlerinin her ikisin birden yapabileceği tezine yol açmıştır. Gerçekten kendini yavaş olarak replike eden sentetik bir ribozim oluşturulmuş ve doğal olarak bilinen intron bağlayıcı üç RNA’nın bir alt birimi de kendi kopyasını oluşturabilme yeteneğindedir. Kimyasal yaklaşım, büyük bir olasılıkla ilk katalizör/gen kombinasyonlarının yalnızca RNA analogları olabileceğini göstermektedir. Tam RNA temelli bir sistem, ikinci kez, kendisinden daha az yeterli bir atasının yerine geçebilir. Yoğun doğal seçilim, daha az yararlı replikasyonları ve metabolik kontrol sistemlerini elemek yönünde işleyecek ve yüksek mutasyon hızına sahip olan bu ilkel formlar arasından, tamir yetenekleri olan sistemleri seçecektir.
Sonraki adım ve teorisiyenler için en zor olan adım translasyonun evrimidir. Enzim ve yapısal elementler olarak, RNA’dan ziyade proteinlerin kullanılmasının canlı sistemlere seçici bir avantaj kazandırması açıkça bilinse dahi, transkripsiyon ve translasyon süreçleri arasında görülen boşluk çok büyük görünmektedir. Nükleik asitlerdeki nükleotit dizileri ile, proteinlerdeki amino asit dizileri arasındaki korelasyonun nasıl ortaya çıktığının bir sır olması gibi, transkripsiyon ile translasyon arasındaki korelasyon da bir sırdır. Ancak, yaklaşık bir tahmini verecek bazı öneriler vardır. İlki, şimdiye kadar bilindiği kadarıyla, genetik kodun hemen hemen evrensel olmasıdır. Çok nadir durumlar hariç, mikroorganizmalardan insana kadar tüm organizmalarda kodonlar aynı amino asitleri şifrelerler. Bu durum, genetik kodun yalnız bir kez ve rastgele ortaya çıkmış olduğu ve bilgi akışının kesilmesine neden olacağından, değişimlerin organizmalar için öldürücü olacağı anlamına gelebilir. Diğer taraftan, hücresel var oluş döneminin tümünde, kodonlarda çok az varyasyonun oluşmuş olduğu, hangi kodunun hangi amino asitlere gittiğinin tamamen rastgele olduğu zorlaması ile çelişebilir. Örneğin, genetik kod sözlüğüne dikkatli olarak bakıldığında, U`nun ikinci harf olduğu tüm kadonların hidrofobik amino asitleri kodladığı ve elektriksel yükü olan 5 (beş) amino asidi kodlayan kodonlarda ortadaki harfi bir pürin (A ya da G) olan kodonlar olduğu görülür. Kısaca, bazı kimyasal sistemler, kodon ve amino asit eşleşmelerinin temelini oluşturmaktadır. Bu kimyasal sistemler için yapılan açıklamada, -şayet kimyasal bir mantığa sahip ise (ki bilinmiyor)- varsayım, kademeli olarak önceki mirasa ekleneceğinden, hem üreme süreci sırasında daha doğru duplikasyonları, hem de damlacıklar içinde meydana gelen kimyasal tepkimeler üzerinde daha kesin bir kontrolü mümkün kıldığı varsayılmaktadır.
 Sonraki adım, özellikle tercih edilen nitelikleri taşıyan canlı öncüllerinin (prebiyontların) küçük bir yüzdesinin yavaş yavaş ilk primitif hücrelere gelişmesi olmuş olmalıdır. “Cansız” prebiyontlardan “canlı” hücrelere ani bir geçişin olmadığının hemen hemen kesin olduğuna dikkat edin. Normalde yaşama atıf ettiğimiz bu özellikler, kademeli olarak kazanılmıştır. Bu evrede, canlı ve cansız arasındaki sınır keyfi (belirsiz) bir sınırdır.
Sonraki adım, özellikle tercih edilen nitelikleri taşıyan canlı öncüllerinin (prebiyontların) küçük bir yüzdesinin yavaş yavaş ilk primitif hücrelere gelişmesi olmuş olmalıdır. “Cansız” prebiyontlardan “canlı” hücrelere ani bir geçişin olmadığının hemen hemen kesin olduğuna dikkat edin. Normalde yaşama atıf ettiğimiz bu özellikler, kademeli olarak kazanılmıştır. Bu evrede, canlı ve cansız arasındaki sınır keyfi (belirsiz) bir sınırdır.
Yukarıda verilmiş olay dizeler, tüm biyologlarca kabul edilmemektedir. Bazıları, ilk canlının nükleik asit gibi kendi kendini replike eden öncüller (çıplak gen) olmasının daha olası olduğunu düşünürler. Bu durumda ilk hücre, bu makromoleküllerin etrafında diğer maddelerin yavaş yavaş birikmesi ile bir kabuğun oluşması sonucu ortaya çıkmış olacaktır. Başka bir deyişle, hücrenin genezisi (ilk ortaya çıkışı) ile ilgili ilk model, önce oldukça ilkel ve gerçek anlamdaki üremeden farklı bir üreme yeteneğine sahip prebiyontların ortaya çıktığını ve sonra yavaş yavaş bir genetik kontrol sisteminin geliştiğini savunur. Fakat bu ikinci model önce kontrol sisteminin ortaya çıktığını ve daha sonra bunun etrafında bir sitoplazma ve zarın geliştiği öne sürmektedir.
Diğer bir senaryo yaşamın kil partikülleri yüzeyi üzerinde geliştiğini varsayar. Kil, belirli molekülleri absorbe etme ve zayıf olarak tepkimeleri katalizleme yeteneğine sahiptir. Daha sonra seçilim en yararlı kimyasal özellikli kimyasal partikülleri tercih edecektir.
Nasıl ortaya çıkmış olursa olsun, canlı hücrelerin bu ilkel öncülleri, kendi çevrelerindeki kimyasal maddeleri besin olarak absorbe etmiş ve aynı zamanda bu kimyasalları replikasyon için gerekli maddelere çevirmeye başlamışlardır. Bunlar “beslenme” ve “üreme” yeteneğinde olan varlıklar olduklarından, artık canlıları tanımlayan kriterlerin en azından bazılarına uymaya başlamışlardır.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç