
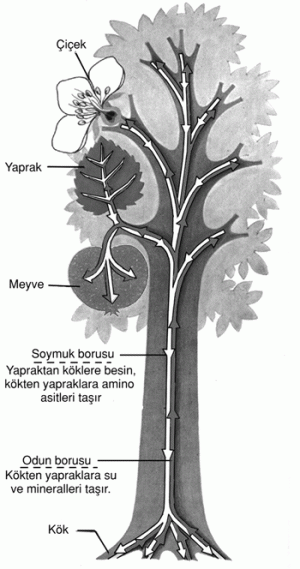
 Özsuyu köklerin absorbladığı su ve suda erimiş elementlerdir. Ksilemdeki gelişmiş trake ve trakeitler de yukarı doğru taşınır. Esas olarak, ksilemde gerçekleşen yukarı yöndeki bu taşınım, halkalama denemeleriyle kolayca gösterilebilir; eğer bir ağacın gövdesinden mantar, flöem ve kambium halka şeklinde çıkarılırsa, yapraklar ile kökler arasında yalnızca ksilem bağlantısı kalsa bile, yapraklar şişkinliklerini konular. Bunun ötesinde, taşınım, olgun ksilemde canlılığını koruyan bir kaç hücreye bağlı değildir. Eğer bu hücreler ısı ya da zehirle öldürülürlerse öz suyunun yükselişi azalmaz.
Özsuyu köklerin absorbladığı su ve suda erimiş elementlerdir. Ksilemdeki gelişmiş trake ve trakeitler de yukarı doğru taşınır. Esas olarak, ksilemde gerçekleşen yukarı yöndeki bu taşınım, halkalama denemeleriyle kolayca gösterilebilir; eğer bir ağacın gövdesinden mantar, flöem ve kambium halka şeklinde çıkarılırsa, yapraklar ile kökler arasında yalnızca ksilem bağlantısı kalsa bile, yapraklar şişkinliklerini konular. Bunun ötesinde, taşınım, olgun ksilemde canlılığını koruyan bir kaç hücreye bağlı değildir. Eğer bu hücreler ısı ya da zehirle öldürülürlerse öz suyunun yükselişi azalmaz.
Sorun
Ksilemde öz suyunun yükselişi için getirilen herhangi bir genellemenin, boyu 90-120 m’ye ulaşan en uzun boylu ağaçların tepe kısımlarına suyun ulaşmasını sağlayan kuvvetleri tanımlaması gerekir. Bu basit bir sorun değildir. Bir tüpü suyla doldurduktan sonra ucunu kapatarak üstü kapatılamamış bir su havuzunun dip kısmına yerleştirdiğimizi düşünelim. Yerçekimi tüpteki suyu aşağı çekecek ve eğer tüp yeterince uzunsa uçta bir vakum oluşacaktır. Bir ağacın ksilemindeki özsuyu buna benzer bir yerçekimi kuvvetinin etkisine maruz kalır.
Tüpteki suyun yüksekliğinin sürekliliği tüpü kuşatan su havuzunun yüzeyindeki hava basıncı tarafından sağlanır. Hava basıncının varlığını hayalimizde canlandırmak pek çoğumuz için kolay değildir; ancak bu basınç tümüyle gerçektir. Üzerimizdeki kilometrelerce kalınlıktaki atmosferin ağırlığı, 1 atmosfer ya da atm olarak gösterilir, deniz seviyesinde, 10.4 m uzunluğundaki bir su sütunun oluşmasını sağlar (Daha yükseklerde sütun büyük ölçüde kısalır). Bu nedenle, en uzun boylu bir ağacın uzunluğuna eşit olan 120 m’lik bir su sütununu desteklemek için en az 12 atm lik bir basınç gereklidir. Fakat sütun bu söylenenden daha fazla olmalıdır. Bu sıvı, bazen dakikada bir metre ya da daha hızlı bir şekilde yukarıya doğru taşınmalı ve bu taşınım, sürtünme kuvveti düz yüzeyli bir tüpten daha fazla olan ksilem gibi bir sistemde gerçekleşmelidir. Bu koşullarda sıvının en uzun ağaçların en yüksekteki dallarına taşınması için en az 30 atm’lik kuvvete gereksinim vardır.
Öyleyse, ksilemde sıvının taşınımına ilişkin herhangi bir kuramın bu büyüklükte bir kuvvetin nasıl oluştuğunu açıklaması gerekir. Bu soru kuşaklar boyu bitki fizyologları arasında sıcak bir tartışma konusu yaratmıştır. Pek çok düşünce ortaya atılmış ve araştırılmıştır. Yıllarca, hiç kimse, özsuyunun aşağıdan mı yukarı itildiği, yoksa yukarıdan mı çekildiği konusunda emin olamamıştır.
Kapillarite (kılcallık)
Uzun bir süre, kapillaritenin bu sıvı hareketinin kaynağı olduğu düşünülmüştür. İlgili kuvvetlerin büyüklüğü konusundaki gerçekçi değerlendirmeler, kapillaritenin özsuyunun yükselmesi gibi muazzam bir olayın açıklanmasında çok yetersiz kaldığını göstermiştir.
Kök Basıncı![Bitkilerde Özsuyunun Taşınması]()

Getirilen diğer bir açıklama kök basıncıdır. Bazı bitki türlerinin gövdeleri kesildiğinde özsuyu belirli bir süre kesik yüzeyden akar. Eğer kesik yüzeye bir tüp oturtulursa bu tüpün içinde bir metre ya da daha uzun bir su sütunu yükselir. Benzer şekilde, koşullar kökler tarafından suyun absorbsiyonu için optimum; ancak havadaki nem, transpirasyonla az su kaybına neden olacak kadar yüksek olduğunda, basınç altındaki su, yaprakların kenarında su damlacıkları oluşturarak yaprak damarlarının uçlarından dışarı atılır. Suyun bu şekilde salgılanması işlemine gutasyon denir. Bu guttasyon ya da sıvı akması örneklerinde olduğu gibi, ksilemdeki sıvı, basınç altında olduğunda, bunu gerçekleştiren itici kuvvetin köklerde bulunduğu açık olup, bu kuvvet kök basıncı olarak adlandırılır. Bu kuvvet gerçekçi bir biçimde ölçülünceye değin, özsuyun akışı kök basıncına dayandırılmıştır.
Kök basıncı nasıl oluşmaktadır? Topraktan alınan suyun kökteki epidermis, korteks, endodermis ve kökü geçerek ksileme, oradan da yukarıya doğru bitkinin diğer kısımlarına taşındığını biliyoruz. Ancak, ksilemde pozitif basınç altındaki uzun bir su sütunu kendi ağırlığına bağlı olarak aşağıya doğru kuvvetli bir hidrostatik kuvvet yaratacaktır. Suyun, bu kuvvet tarafından köklerde ksilemin dışına sürülmesi beklenebilecektir. Halbuki, su, yalnızca kökün merkezi silindirinde tutulmakla kalmayıp, aynı zamanda sütunun yukarıya doğru itilebilen bir kuvveti oluşturabilecek miktarda merkezi silindire taşınmaya devam eder. Ancak kökler öldürüldüğü taktirde kök basıncı tamamen ortadan kalkar ya da kökler oksijensiz bırakılırsa kök basıncı durur. Bu, kök basıncının gelişmesi için solunumla ATP üretiminin gerekli olduğunu göstermekdir.
ATP’den sağlanan enerji ile aktif iyon taşınımını sürdürüldüğü, bunun da azalan bir su potansiyeli oluşturduğu açıktır. Endodermisin dış tarafındaki kök hücrelerindeki iyonlar iç tarafa pompalanırlarken perisikldeki iyonlar ksileme pompalanırlar. Sonuç olarak, simplast yolunu izleyen su, endodermisten köklere ve ksilemdeki trakelere çekilir. Apoplastta ilerleyen su ise endodermisteki düşük su potansiyeli tarafından, su potansiyeli endodermisinkinden düşük olan ksileme çekilir. Ancak, kapilarite gibi kök basıncı da sorunun tam ya da hatta bu büyük bir kısmına yanıt veremez. Kök basıncının ortaya çıktığı türlerde, bu basıncın seyrek olarak 1 ya da 1 atm’i aştığı saptanmıştır.
Birçok araştırmacı tarafından kök basıncı değerlerinin düşük bulunmuş olması, en azından uzun boylu ağaçlarda özsuyunun yükselmesinde ana itici kuvvetin kök basıncı olduğundan kuşku duyulmasının tek nedeni değildir. Eğer yaz aylarında ksilemde bir delik açılırsa, su, basınç altında kalmış gibi fışkırmaz. Bunun aksine, trakelerin içine hava çekildiğini anımsatan kısa bir ıslık sesi oluşabilir. Halbuki, yazın büyük miktarlardaki su yukarı doğru taşınır.
Özetle, su, düşük bir su potansiyel gradienti yönünde taşınabilir; ancak oluşan kök basıncı gövdede özsuyunun nasıl taşındığını açıklamaya yetmez. Kök basıncının, olsa olsa, bazı bitkilerde, özellikle de ilkbahar başlangıcında, çok genç bitkilerde, özsuyunun yukarıya doğru taşınmasında rolü olabilir.
TATC Teorisi
Yapraklarda, parankima hücrelerinin çeperlerindeki (ya da gövdenin diğer kısımlarında) su, buharlaşmayla kaybedildikçe, onun yerini hücre içeriğindeki suyun aldığını biliyoruz. Suyun uzaklaşması sonucu yaprak hücrelerinin su potansiyelleri düşer ve bunlar komşu hücrelerden, bu hücreler de onlara komşu hücrelerden suyu çekerler. Bu şekilde, yaprak damarlarındaki ksileme doğru bir su potansiyeli gradienti oluşur. Ksilemdeki su potansiyeli gradienti, ksileme bitişik parankima hücrelerinin ksilemdeki suyu çekebilmelerini sağlar. Bu işlem, su sütununun yukarı doğru çekilmesine yardım eder. Burada su sütunun hava basıncı ya da vakumla yukarı çekilmediği göz önünde bulundurulmalıdır; bu mekanizma bir tahıl sapı tarafından bir sıvının emilmesine benzemez. Hava basıncı suyun yalnızca 10.4 m yükselmesini sağlar. Bu durumda, hücre çeperinin buharlaşma yüzeylerindeki su ile ksilemdeki ve köklerdeki su ile en uçtaki ksilemdeki su arasında bir süreklilik gerekmektedir. Sistemin içine hava girmesi nedeniyle yaprak hücrelerinden köke kadar olan bu süreklilik bozulduğu taktirde, bu özel ksilem yolu işlevini durduracaktır.
Başlıca buharlaşmanın bir sonucu olarak suyun yukardan çekildiğine ilişkin görüş, buharlaşmayı (ya da onun değişiyle “perspirasyon”) keşfeden öncü botanikçi, İngiliz Papaz Stephan Hales tarafından 18. yüzyılın başlarında ortaya atılmıştır. 1984 yılında ise İrlandalı Botanikçi H.H. Dixon ve onun fizikçi arkadaşı J.Joly tarafından daha ayrıntılı bilgiler verilmiştir. Bu görüş, her bir su molekülü arasında çok büyük bir kohezyon bulunmasına ve bu su moleküllerinin ksilem borularına ve yapraklardaki mezofil hücrelerinin hidrofilik çeperlerine tutunma eğilimlerine dayandırılmaktadır. Ksilemde yukarı doğru taşınan su moleküllerinin arkadaki diğer su moleküllerini çekmeleri gerekir. Burada, su molekülleri arasında ya da hücre özsuyu ve trakelerin çeperleri ve daha sonra da mezofil hücrelerinin çeperleri arasında hiçbir kopukluk ve ayrılma olamaz.
Su molekülleri, hidrojen bağları nedeniyle gerçekten büyük bir kohezyon gösterirler. Teoride, bütün bir su sütununun kohezyon kuvveti (köklerden yapraklardaki transpirasyon yüzeylerine kadar uzanan, çok dar ve çok hidrofobik alan borularla kuşatılmış hidrojen bağlı moleküllerin oluşturduğu kesintisiz bir sistem) 15.000 atm ye kadar çıkabilir. Gerçek deneysel değerler daha düşük olmasına karşın, bu değer 300 atm. ya da daha fazla olabilir. Eğer hücrede özsuyun yükselmesi 30 atm. ya da daha fazla bir basınç gerektirirse, bu değerler Dixon ve Joly’nin görüşüne uygunluk gösterir. Bu görüş genel olarak transpirasyon-adezyon-tansiyön- kohezyon (TATC) teorisi ya da basitçe transpirasyon teorisi olarak adlandırılır, . (“Adezyon”, suyun, ksilem elemanlarının ve çok büyük bir yüzey alanı oluşturan yaprak mezofil hücrelerinin çeperlerine bağlanmasını ifade eder; “tansiyon” ksilemdeki suyun su potansiyeli düşük olan yaprakların içine çekilmesidir; “Kohezyon” ise ksilem sütununda ve mezofildeki su molekülleri arasındaki hidrojen bağlandır).
Transpirasyon sisteminin enerji kaynağı güneştir. Güneş ksilemden suyun yukarı çekilmesini sağlar.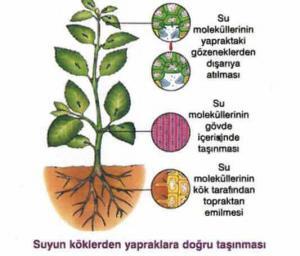
TATC teorisi çok uzun boylu ağaçların sahip oldukları koşullarda denenmemiş olmakla birlikte, 1890’da Avusturya’da Jozef Böhm ve Almanya’da E. Askenasy daha küçük ölçekli bazı ilginç denemeler gerçekleştirmişlerdir. Bu araştırıcılar, alt ucu civaya daldırılmış çok ince bir cam tüpün üst kısmından suyun buharlaşması halinde, buharlaşmaya bağlı olarak, civa sütununun tüpün üst kısımlarına doğru çekileceğini göstermişlerdir. Eğer kesik bir dalın alt kısmı ince bir borunun üst ucuna sıkıca yerleştirilir ve bu daldaki yapraklarda buharlaşma meydana gelirse, benzer olay görülür. Su moleküllerinin ağır bir civa sütununun yukarı çekilmesine yetecek kadar birbirlerine (ve tüpün çeperlerine) sıkıca yapıştıkları açıktır.
TATC teorisine başka denemelerden de destek gelmektedir. 1935’te Alman Botanikçi Bruna Huber ksilemin içine küçük elektrik cisimleri yerleştirerek özsuyunu ısıtmıştır. Araştırıcı daha sonra, ağaç üzerinde, özsuyunun biraz üzerine bir ısı ölçer yerleştirerek ısınmış özsuyunun pilden geçiş süresini ölçmüştür. Huber, sabahları, suyun, ağacın üst kısımların da gövdenin alt kısımlarından daha önce taşınmaya başladığını bulmuştur. Bu, özsuyunun yukarı yöndeki hareketinin ağacın kaidesinde değil, ağacın üst kısmında başladığının göstermektedir. Günün farklı saatlerinde ağaç gövdelerinin çevresinin titizlikle ölçülmesi sonucu daha fazla kanıt elde edilebilir. Transpirasyon hızının yüksek olması nedeniyle ksilemdeki gerilimin en üst düzeye çıktığı gündüz saatlerinde gövdenin çevresi küçülür-yani, kapalı, esneyebilir, içi sıvı dolu herhangi bir tankın içeriğinin aynen yukarıya çekilmesinde olduğu gibi gövdenin çapı azalır. Yukarıya doğru çekilmenin nedeni buharlaşmadır.
Bitkilerin çoğunda ksilemde yükselen suyun % 90’ının, fotosentez ya da büyüme için tüketilmeden buharlaşmayla kaybolduğu gösterilmiştir.
TATC teorisi bitki fizyologları arasında geniş kabul görmüş olmasına karşın, geride bir dizi çözümlenmemiş soru bırakmıştır. Bu teori, ksilemdeki su sütununun hiç aralıksız sürdürülmesine gereksinim gösterir; halbuki, sütunda sıklıkla kopmalar oluşmaktadır. Örneğin, kuraklık sırasında özsuyunun içinde çözünmüş bazı gazlar, gaz kabarcıkları oluşturabilirler ve yine özsuyu, kışın donduğunda, çözünmüş gazlar su sütunu parçalayan kabarcıklar oluşturarak baskı yaparlar.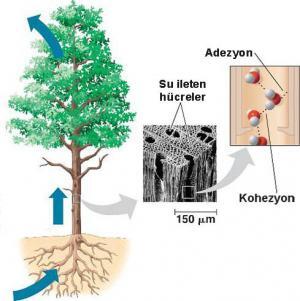
Su sütunlarındaki bu kopmalar sonucu, ksilem elemanlarının daha büyük bir bölümü giderek etkilendikçe, bir gövdenin toplam iletkenliği azalır. Bitkiler bu hava kabarcıklarının onarılması ya da izolasyonu için bazı stratejiler geliştirmişlerdir. Örneğin otsu bitkilerde gece oluşan kök basıncı (trakelerde gerilim azaldığında), kırılan su sütunlarını tekrar birleştirebilir. Donma sırasında çözeltiyi sıkıştaran gazlar sıcaklık arttığında tekrar çözünebilirler. Buna ek olarak, trakeitlerdeki geçitler, sıvıların ksilemin gazla dolmuş ya da zarar görmüş kısımlarına bypass ile geçişine izin verirler. Ayrıca gövdede onarılamayan sütunlardan herhangi birinin yerini yeni sentezlenmiş ksilem halkaları, alır.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç