
 İlk bakışta eşey, evrimsel bir anlam vermemektedir. Bir dişinin üreme çabalarının yarısı döl vermeyen ve dolayısıyla o dişinin üreme verimine çok az katkıda bulunan erkek döllerinde devam eder. Eğer erkekler elimine edilebilirse, her bir dişi üretebileceği dişi döllerinin iki katı kadar dişi ve onlardan çok daha fazla torun dişi verebilecektir. Bu hem eşeyli hem de eşeysiz formlara sahip yüksek organizasyonlu bitki ve hayvan gruplarında kolayca gösterilebilir. Yukarıda değinilen bu organizmaların hemen tümünde, eşeysiz formlar daha sonra eşeyli çoğalan öncüllerden ortaya çıkmıştır.
İlk bakışta eşey, evrimsel bir anlam vermemektedir. Bir dişinin üreme çabalarının yarısı döl vermeyen ve dolayısıyla o dişinin üreme verimine çok az katkıda bulunan erkek döllerinde devam eder. Eğer erkekler elimine edilebilirse, her bir dişi üretebileceği dişi döllerinin iki katı kadar dişi ve onlardan çok daha fazla torun dişi verebilecektir. Bu hem eşeyli hem de eşeysiz formlara sahip yüksek organizasyonlu bitki ve hayvan gruplarında kolayca gösterilebilir. Yukarıda değinilen bu organizmaların hemen tümünde, eşeysiz formlar daha sonra eşeyli çoğalan öncüllerden ortaya çıkmıştır.
Azalan üreme verimine ek olarak, eşeylilik üreme başarısında etkin olarak işe karışır. Bir organizmanın üremesini sağlayan ve yeterince işleyen bir alel takımı her bir gamet için rasgele olmak üzere yarı yarıya bölünür. Bu aynen tam bir deste iskambil kağıdı ve tümü tek renkli ful bir elden rasgele kartların çekilmesi ve yeni bir el oluşturulması demektir. Bu yeni iskambil eli, ilk poker eli kadar güzel olmayacaktır. Bazı eşeysiz türler daha başarılıdır; hemen tüm karahindibalarda çiçekler olmasına karşın eşeysel üreme yeteneğinin olmaması gibi.
Eşeyin evrimi, tahminen normalde eşeysiz olarak çoğalan bakteri gibi haployit eşeysiz türlerle başlamıştır. Yeterli besin bulunduğunda, Escherichia coli 20 dakikada bir üreyebilir. Kromozomun replikasyonu 18 dakika sürdüğünden, koşullar uygun olduğu sürece döl verme mücadelesi zamana karşı olan bir yarıştır.
Diployitlik, eşeyliliğe doğru sonraki adımdır ve kromozomunun replike edeceği iki kopyası olan bir bakterinin önemli bir üreme dezavantajına sahip olması açıktır. Bu bakteri haployit durumunda iki saatte 64 döl verirken diployit durumda sadece 8 döl verecektir. Ancak, haployidinin dezavantajı herhangi bir mutasyonun hemen ifade edilmelidir ve çoğu genetik değişimler zararlı olduklarından, bunun dezavantajı daha yüksek olabilir. Sonraki bölümlerde göreceğimiz gibi, bu durum bitki ve hayvanlardaki benzer modellerde görülen diployit baskınlığın evrimini açıklamak için en olası faktördür. Bunlarda mutasyonal tamir aktivitelerine harcanan kaynaklar ömür uzunluğu ile artar.
Diployitlik tek başına eşeysel rekombinasyon demek değildir; (karahindibalar diployittir)  ancak, eşeysel rekombinasyon olmaksızın bile bir çok avantaj sağlar. En açık şekli ile, zararlı mutasyonlar gizlenir. Ayrıca, tam bir kromozomda, iki zincirden biri diğerinde olan DNA hasarının tamirinde kalıp olarak kullanılabilir. Aynı zamanda, diployitler, koşullar değiştiğinde yararlılığı denebilecek olan varyantları yedek kopyalarında saklayabilirler.
ancak, eşeysel rekombinasyon olmaksızın bile bir çok avantaj sağlar. En açık şekli ile, zararlı mutasyonlar gizlenir. Ayrıca, tam bir kromozomda, iki zincirden biri diğerinde olan DNA hasarının tamirinde kalıp olarak kullanılabilir. Aynı zamanda, diployitler, koşullar değiştiğinde yararlılığı denebilecek olan varyantları yedek kopyalarında saklayabilirler.
Eşeysel üreme, krossing-over ve genetik rekombinasyon sağlar. Krossing-overde, eşeyli diployitlerin DNA tamiri için kullanılan ve bakterilerin konjugasyonları sırasında nadiren kullanılan aynı tip enzimler işlev gördüğünden, öyle anlaşılıyor ki, olası ilk eşeysel tür, gamet oluşumundan önce ortaya çıkan krossing-overin hataları düzeltme avantajlarından yararlanmışlardır. Fakat değişiklikler, yeni olanaklar sağlamış ve biyologların çoğu, krossing-overin tamir ettiği hatalardan çok yeni varyasyonlar yarattığı konusunda hem fikirdirler.
Krossing-over ve rekombinasyonun evrimdeki rolü ile ilgili çalışmalar, varyasyonun rolüne ilişkin iki genel hipotezi doğurmuştur. Tangled-bank (rastgele yıgılma) modeli olarak isimlendirilen ilki, The Origin of Species’in hemen sonuna yakın verilen, çok sayıda tür içeren ve değişik koşulları olan bir ortam modeli üzerine kurulmuştur. Darwin, koşulların 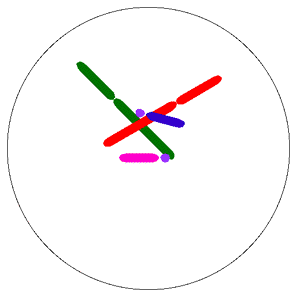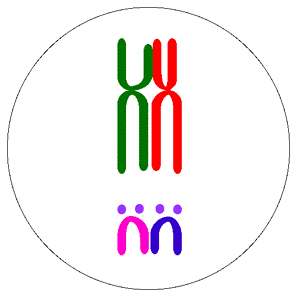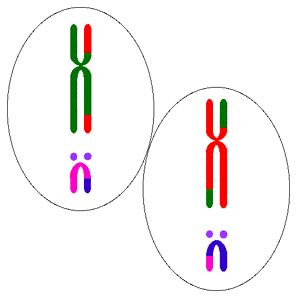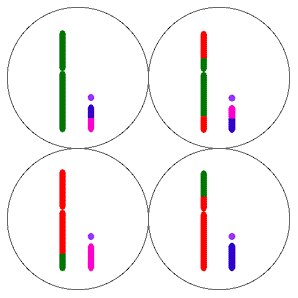 aynı olduğu her yerde bir kaç türün baskın olacağını varsaymıştır. Ancak, kendisi 1 m^2 çayırlığını incelediğinde 20’den fazla bitki türü saymıştır. Darwin’in rastgele yığın tanımlaması, küçük bir alanda bile bulunan çok sayıda farklı mikrohabitatın önemini vurgulamıştır. Rastgele yığılma (tangled bank) modeline göre, eşeysiz üreyen türler, eşeyli üreyen türlere göre dezavantajlıdır. Çünkü, eşeyli üreyenler mikrohabitatlara göre bir dizi döl verebilirken eşeysiz üreyen türler yalnız bir habitatta üstün olacaklardır. Uyum sınırları dar olduğundan, eşeysiz kardeşler, her şeyi aşmış olsalar bile, esas olarak birbirleri ile rekabet edeceklerdir. Halbuki, bir çiftin birbirine benzemeyen eşeyli dölleri kendine en uygun yerleri bulmak için yeterince farklı olmalıdır. En azından bazı türlerden bu hipotez için iyi kanıtlar sağlanmıştır.
aynı olduğu her yerde bir kaç türün baskın olacağını varsaymıştır. Ancak, kendisi 1 m^2 çayırlığını incelediğinde 20’den fazla bitki türü saymıştır. Darwin’in rastgele yığın tanımlaması, küçük bir alanda bile bulunan çok sayıda farklı mikrohabitatın önemini vurgulamıştır. Rastgele yığılma (tangled bank) modeline göre, eşeysiz üreyen türler, eşeyli üreyen türlere göre dezavantajlıdır. Çünkü, eşeyli üreyenler mikrohabitatlara göre bir dizi döl verebilirken eşeysiz üreyen türler yalnız bir habitatta üstün olacaklardır. Uyum sınırları dar olduğundan, eşeysiz kardeşler, her şeyi aşmış olsalar bile, esas olarak birbirleri ile rekabet edeceklerdir. Halbuki, bir çiftin birbirine benzemeyen eşeyli dölleri kendine en uygun yerleri bulmak için yeterince farklı olmalıdır. En azından bazı türlerden bu hipotez için iyi kanıtlar sağlanmıştır.
İkinci hipotez, Through the Looking Glass(ters yönlü oluk) ‘dan esinlenen red-queen modelidir. Through the looking Glass ‘da, Alis ve Kızıl Kraliçe koşabildikleri en yüksek hızla koşarlar; ancak ne kadar hızlı koşarlarsa koşsunlar herhangi bir gelişme kayıt edemezler ve bulundukları alanın dışına çıkmazlar. Red queen kuramı, varyasyon için duyulan gereksinimin koşullardaki değişim oranında, predatör, avcı ve parazit zeminini hızlı bir evrimleşme ile koruma ihtiyacından kaynaklandığını savunur. Buradaki vurgulama, alandaki varyasyondan ziyade zamandaki varyasyondadır. Bu açıklama, özellikle parazitlerin durumu ile daha iyi anlaşılır: çoğu parazitler konak soyuna özgül olduklarından, bunların eşeysiz üreyen bir populasyonu tamamen ortadan kalkabilir. Örneğin İngiltere’de, 3-5 yıl kadar 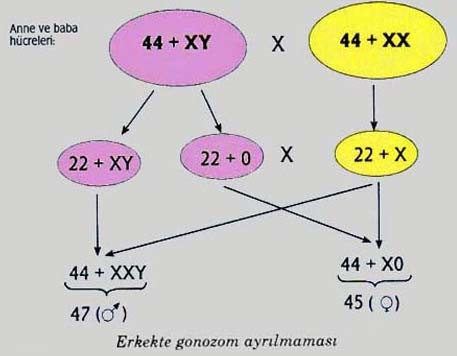 önce, ürünleri harap eden fungal bir parazit bulaşmadan önce, bir arpa soyu normal bir şekilde yetişebilmekteydi. Uzun ömürlü konaklar üzerinde yaşayan tripslerle (Thysanoptera = bitki bitleri) yapılan çalışmalar, zamanla bir bitkiye özelleşen kolonilerin geliştiğini göstermiştir. Parazitler, konakları ile karşılaştırıldığında daha kısa bir döl verme zamanına sahip olduklarından bu özelleşme olasıdır. Konak bir defa üreyene kadar parazit çok sayıda seçilimden geçebilir.
önce, ürünleri harap eden fungal bir parazit bulaşmadan önce, bir arpa soyu normal bir şekilde yetişebilmekteydi. Uzun ömürlü konaklar üzerinde yaşayan tripslerle (Thysanoptera = bitki bitleri) yapılan çalışmalar, zamanla bir bitkiye özelleşen kolonilerin geliştiğini göstermiştir. Parazitler, konakları ile karşılaştırıldığında daha kısa bir döl verme zamanına sahip olduklarından bu özelleşme olasıdır. Konak bir defa üreyene kadar parazit çok sayıda seçilimden geçebilir.
Red queen hipotezinin işaret ettiği noktalardan biri, krossingoverin en uzun yaşayan türler arasında daha yaygın olması gerektiğidir. Bu durum, konaklara, parazitin hızlı evrimleşmesini dengelemek için, mümkün olduğunca farklı döller oluşturarak bir telafi yeteneği kazandırmaktadır. Ömür uzunluğu ile krossing-over hızının artışı, en azından memelilerde ki modelde görülebilir. Anlaşılması zor olan şey, parazitlerin karahindiba ve diğer eşeysiz organizmaları neden ortadan kaldırmadığıdır.
Her iki hipotez, habitat varyasyonu konusunda, hem eşeyli hem de eşeysiz üreyebilen bir çok türde görülen modellerle uyuşmaktadır. Örneğin, uygun yeni bir bitki sürgünü üzerine yerleşen ilk yaprak biti hızla eşeysiz olarak üremeye başlar ve bu lokal habitata iyi bir şekilde uyum gösteren bir dişi kolonisi oluşturur. Ancak, koşullar kötüleştiğinde, (sürgün üzerindeki yaprak bitlerinin aşırı çoğalması ya da kıtlığa tahammül edememe gibi) kurucular (ilk yerleşenler) kanatlı ve eşeyli olarak üreyebilen döller vermeye başlar. Bu eşeyli bireyler, diğer afidlerle çiftleşir ve uygun bir konak aramaya başlarlar. Açıkça, yaprak biti yaşam döngüsü, tahmin edilemeyen devamlı ya da geçici bir varyasyon olarak açıklanabilir. Eşeyli üreme avantajının derecesi ve doğasının türler arasında farklı olabileceğine inanmak için bir çok neden vardır. Ancak, genellikle, eşeyliliğin varyasyon yarattığı için seçildiği görüşü yaygındır. Mutasyonlarla olandan daha hızlı bir değişim kapasitesi olmasaydı, uzun ömürlü türler yok olurdu.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç