
 Evrim, birbirine benzemeyen çok sayıda türün ortak bir ataya sahip olduğuna ve tüm yaşam formlarının büyük bir olasılıkla aynı uzaklıktaki bir başlangıçtan köken aldığına işaret etmektedir. Zaten evrimin biyologlara yüklediği görevlerden biri, bugün yaşayan türler arasındaki akrabalıkları ortaya çıkarmak ve türedikleri ataları bulmaktır.
Evrim, birbirine benzemeyen çok sayıda türün ortak bir ataya sahip olduğuna ve tüm yaşam formlarının büyük bir olasılıkla aynı uzaklıktaki bir başlangıçtan köken aldığına işaret etmektedir. Zaten evrimin biyologlara yüklediği görevlerden biri, bugün yaşayan türler arasındaki akrabalıkları ortaya çıkarmak ve türedikleri ataları bulmaktır.
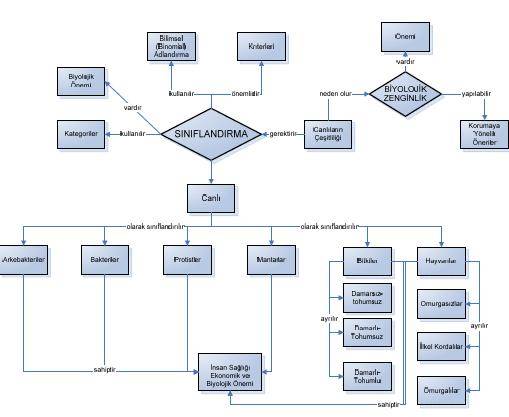
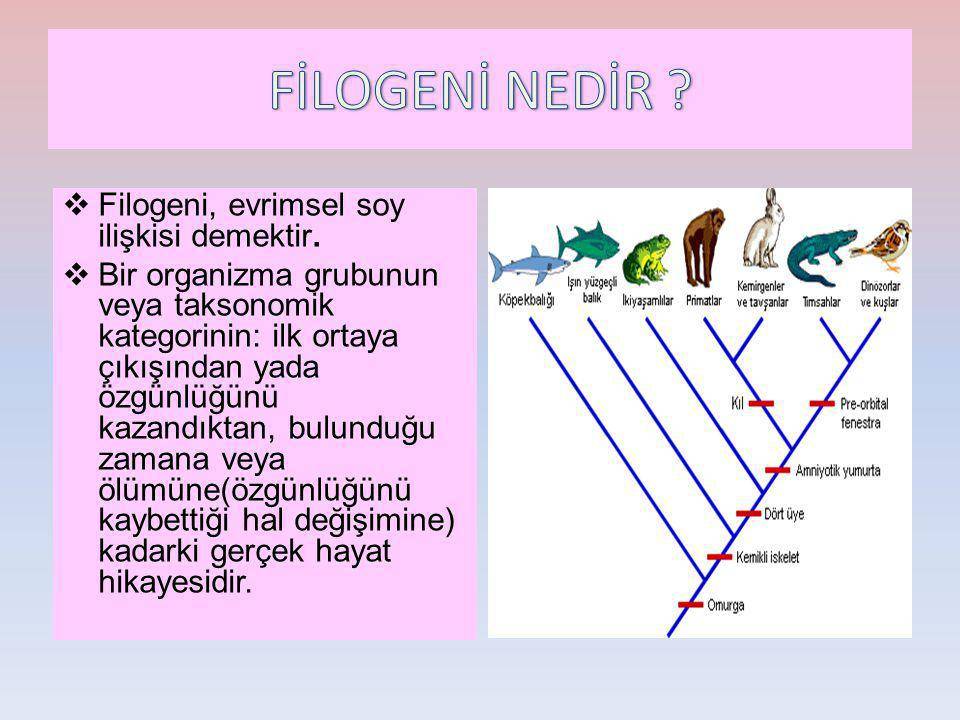
Filogenetik Akrabalıklar
Taksonomist olarak da bilinen sistematikçiler, akraba olduklarını düşündükleri bir grup türün evrimsel geçmişlerini -filogeni- yeniden oluşturmaya (saptamaya) koyulduklarında bugün yaşayan türlerden ve fosil kayıtlardan daha öncesinin olduğunu görürler.
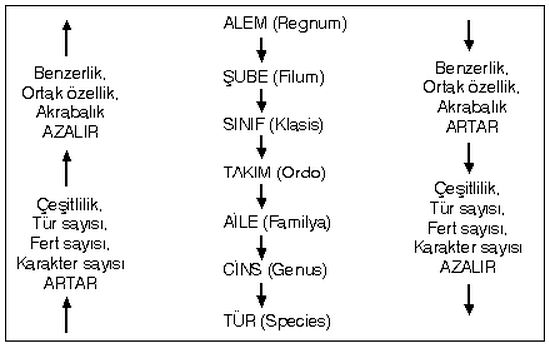
Doğruya en yakın filogenetik öyküyü oluşturmak için bu araştırmacıların gözlemsel ve deneysel verilere dayalı çıkarsamalar yapmaları gerekir. Amacın akrabalık belirlemek olmasına karşın neyin benzerlik olarak dikkate alınacağı zorluk yaratır. Günümüzde 4 temel sistematik yaklaşım vardır. Klasik evrimsel taksonomi, fenetik, kıladistik ve moleküler taksonomi. Her biri benzerliklerden akrabalıkları çıkarmak için farklı teknikler kullanır.
Klasik Evrimsel Taksonomi
Klasik sistematik, diğer yaklaşımlara göre daha fazla tecrübeye ve subjektif yargıya dayalıdır. Klasik yöntemle filogeni oluşturmadaki alışılmış işlem, mümkün olduğu kadar çalışılan türün çok sayıda karakterini çalışmak ve türlerin hangi karakterler farklı, hangi karakterler açısından ise benzer olduklarını belirlemektir. Böylece kısmi farklılık ve benzerliklerin en azından grubun doğru filogenetik akrabalığını kısmen yansıtacağı varsayılır. Genellikle herhangi bir karakterden elde edilecek yanlış verilerinin diğer karakter setinden edinilecek verilerle saptanabileceği düşünülerek, mümkün olan en fazla sayıda farklı karakter kullanılır.
Morfoloji -dış morfoloji, iç anatomi ve histoloji (doku tipleri) ve hücre çekirdeğindeki kromozom morfolojisi- en kolay çalışılan ve yaygın olarak kullanılan karakterlerdir. Yaşayan türlerin karakterleri, bunların fosil formları ile karşılaştırılabildiğinde doğal olarak yararlıdır. Karşılaştırma için seçilen karakterler, analiz edilmekte olan grup içinde değişkenlik gösterenler olmalıdır. Gruba özgü, dolayısıyla ortak ve nispeten yeni bir evrimsel orijine 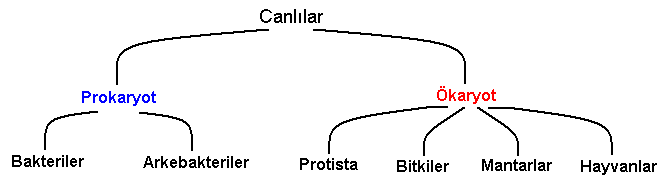 sahip karakterler özellikle kullanışlıdır. Örneğin, Darwin’in ispinozları arasında fosillerde de incelenebilen morfolojik karakterler gaga uzunluğu, gaganın uzunluğunun genişliğine oranı, gaganın başa göre oransal açısı, kafatasını oluşturan farklı kemiklerin lineer uzunluğu, kafatası kemiklerinin göreceli alanı, kasların ses kutusuna bağlanma modelleri (kemiklerde görülen küçük girintilerden) ve benzerleridir. Fosillerden kaybolmuş, ancak yaşayan formlardan ölçülebilen karakterler ise kuyruk uzunluğu, gençlerin desenlenmesi (örneğin işaretin derecesi), desenlenmenin yaşla değişimi ve benzeri karakterleri kapsar. Bunlardan gaga büyüklüğünün çeşitli oluşu, gaga açısının daralması ve nispeten kısa kuyruklar, Darwin’in ispinozlarını, Güney Amerika ana karasındaki kuzenlerinden ayırır.
sahip karakterler özellikle kullanışlıdır. Örneğin, Darwin’in ispinozları arasında fosillerde de incelenebilen morfolojik karakterler gaga uzunluğu, gaganın uzunluğunun genişliğine oranı, gaganın başa göre oransal açısı, kafatasını oluşturan farklı kemiklerin lineer uzunluğu, kafatası kemiklerinin göreceli alanı, kasların ses kutusuna bağlanma modelleri (kemiklerde görülen küçük girintilerden) ve benzerleridir. Fosillerden kaybolmuş, ancak yaşayan formlardan ölçülebilen karakterler ise kuyruk uzunluğu, gençlerin desenlenmesi (örneğin işaretin derecesi), desenlenmenin yaşla değişimi ve benzeri karakterleri kapsar. Bunlardan gaga büyüklüğünün çeşitli oluşu, gaga açısının daralması ve nispeten kısa kuyruklar, Darwin’in ispinozlarını, Güney Amerika ana karasındaki kuzenlerinden ayırır.
Fosillerde korunmuş karakterler özel bir öneme sahiptirler. Çünkü fosil kayıtları eski formların geçirmiş olduğu evrelerle kanıtların doğrudan kaynağıdır. Maalesef fosil kayıtlar tam değildir ve birçok organizma grubu için ise hiç uygun kayıt yoktur.
En iyi açıdan fosiller büyük grupların evrimine genel bir model oluşturur. Bazı gruplarda örneğin çok iyi bilinen atlar gibi, fosil kayıtları, diğer kaynaklardan edinilemeyecek kadar fazla filogenetik bilgi verirler.
Sıklıkla kullanılan diğer bir veri kaynağı embriyolojidir. Eğer morfolojik özelliklerin gelişim seyirleri biliniyorsa filogeni için kullanmak daha kolaydır. Örneğin, A organizmasındaki belirli bir yapı ile B organizmasındaki oldukça farklı görünüme sahip bir yapının her ikisi de aynı embriyonik kökenden (primordiyadan) gelişirlerse A ve B organizmalarının bu yapılarında görülen benzerlik ve farklılıklar, iki organizmanın filogenileri hakkında bilgi verir. Eğer bunlar tamamen farklı embriyonik yapılardan gelişiyorlarsa çıkarım farklı olacaktır.
 Evrimsel kanıtlar, biyologlara çoğunlukla önemli yapılarda ortaya çıkmış olası evrimsel değişimleri izleme imkanı verir ve günümüz yaşam formlarının oluşumuna yol açan olası evrimsel olaylar zincirini kurgulamada yardımcı olur. Örneğin, insanlar dahil memelilerin erken embriyonik gelişimi sırasında faranjiyal (yutak) solungaç keselerinin gelişiminin, karasal omurgalıların uzak atalarının sucul olduğunu gösterdiği düşünülür.
Evrimsel kanıtlar, biyologlara çoğunlukla önemli yapılarda ortaya çıkmış olası evrimsel değişimleri izleme imkanı verir ve günümüz yaşam formlarının oluşumuna yol açan olası evrimsel olaylar zincirini kurgulamada yardımcı olur. Örneğin, insanlar dahil memelilerin erken embriyonik gelişimi sırasında faranjiyal (yutak) solungaç keselerinin gelişiminin, karasal omurgalıların uzak atalarının sucul olduğunu gösterdiği düşünülür.
Yaşam öyküleri klasik filogenetik çalışmalarda önemli rol oynamıştır. Örneğin algleri ve damarlı bitkileri incelediğimizde göreceğimiz gibi bitkilerin yaşam döngüleri sırasında geçirdikleri evreler özellikle önemli bir bilgi kaynaklarıdır.
Konvergens Sorunu
Öyleyse klasik yaklaşım, bir dizi özellikteki benzerliklerin değerlendirilmesine dayalıdır. Fakat ortak evrimsel geçmişe işaret etmek için yalnız benzerlikler yeterli değildir. Belirli bir benzerlik, aynı çevresel koşullara benzer uyumların gösterilmesi sonucu oluşabilir. Bu son durum doğada yaygındır ve filogenetik çalışmalarda önemli bir karışıklık nedenidir. Yakın akraba olmayan organizmalar, benzer çevresel durumlara adaptasyon nedeniyle bir ya da daha fazla karakter açısından daha benzer hale geldiklerinde bunların konvergent evrim (daralan evrim) geçirmiş oldukları söylenir ve bu durum konvergens olarak adlandırılır. Karasal memeli atalardan türemiş olan balinalar, atalarının pergel şeklindeki ayaklarından gelişmiş yassı yüzgeçsi üyelere sahiptir, bu üyeler yüzeysel olarak balıkların yüzgeçlerine benzerler, fakat bu benzerlik konvergensten kaynaklanır ve balina ve balıklar arasında yakın bir akrabalığı göstermez.
Hem eklembacaklılar hem de karasal omurgalılar eklemli üyelere ve hareketli çenelere 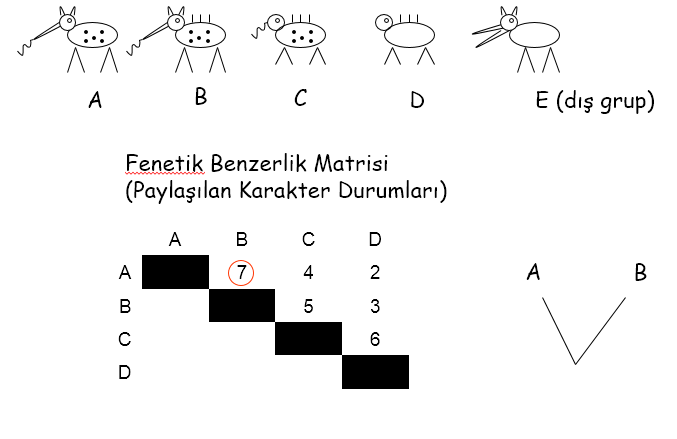 sahiptirler, fakat bu benzerlikler eklembacaklıların ve omurgahların eklemli üye ve hareketli bir çeneye sahip ortak bir atadan evrimleştiğine işaret etmez, bu iki hayvan grubunun eklemli üye ve hareketli çeneleri, üyesiz bir atadan bağımsız olarak evrimleşmişlerdir. Avustralya kör fareleri gerçek kör fareler olmayıp, keseli farelerdir (Yavrusu embriyonik gelişimin erken evrelerinde doğan ve gelişimini bir plasenta için de değil de ananın karın kısmında bulunan bir kese içinde tamamlayan memeli hayvanlardır). Bu keseli fareler, Avustralya’da gerçek kör farelerin dünyanın diğer bölgelerinde paylaştıkları habitatlarla aynı niteliklerdeki habitatlarda yaşarlar, bunun sonucu olarak konvergent olarak evrimleşmiş gerçek kör fareleri andıran bir şaşırtıcı benzerliğine sahiptirler. Ancak keseli kör fareler, diğer kıtalarda yaşayan plesantalı memelilerle, belirgin olarak konvergent olan Avustralya keselilerinin büyük bir grubunu oluşturmaktadır.
sahiptirler, fakat bu benzerlikler eklembacaklıların ve omurgahların eklemli üye ve hareketli bir çeneye sahip ortak bir atadan evrimleştiğine işaret etmez, bu iki hayvan grubunun eklemli üye ve hareketli çeneleri, üyesiz bir atadan bağımsız olarak evrimleşmişlerdir. Avustralya kör fareleri gerçek kör fareler olmayıp, keseli farelerdir (Yavrusu embriyonik gelişimin erken evrelerinde doğan ve gelişimini bir plasenta için de değil de ananın karın kısmında bulunan bir kese içinde tamamlayan memeli hayvanlardır). Bu keseli fareler, Avustralya’da gerçek kör farelerin dünyanın diğer bölgelerinde paylaştıkları habitatlarla aynı niteliklerdeki habitatlarda yaşarlar, bunun sonucu olarak konvergent olarak evrimleşmiş gerçek kör fareleri andıran bir şaşırtıcı benzerliğine sahiptirler. Ancak keseli kör fareler, diğer kıtalarda yaşayan plesantalı memelilerle, belirgin olarak konvergent olan Avustralya keselilerinin büyük bir grubunu oluşturmaktadır.
Bu tartışmalar, klasik sistematikçiler iki tür arasında benzerlikler bulduklarında, bu benzerliklerin olasılıkla homoloji mi (ortak bir atadan kalıtılan) yoksa sadece anoloji mi (işlev ve çoğunlukla yüzeysel yapıda benzer; ancak evrimsel orijinleri farklı ) olduğunu belirlemeye çalışması gerektiğini göstermektedir. Böylece kanıtlar, her iki kuşun kanatlarının ortak bir kuş atanın kanatlarından geliştiğine işaret ettiğinden ardıç kuşu ve mavi kuşun kanatlarının homolog olduğu düşünülmektedir. Fakat ardıç kuşunun kanatları ile kelebeğin kanatları, işlevsel olarak benzer yapılar olmalarına karşın yalnızca analogturlar. Çünkü bunlar, ortak bir atadan kalıtılmayıp bağımsız olarak evrimleşmişler ve farklı atasal yapılardan oluşmuşlardır.
İki yapının ne anlamda homolog olduğu ya da anolog olarak kabul edileceğine işaret edilmesi her zaman önemlidir. Örneğin, kuşların ve yarasaların kanatları bağımsız olarak geliştiğinden homolog değillerdir, ancak her ikisinin kanatları homolog kemikler içerirler. Kuş ve yarasanın kanatları kuş ve memelilerin atası olan eski bir karasal omurgalının ön üyelerinden evrimleşmişlerdir. Kısacası, yarasa ve kuşların kanatları kanat olarak anolog, ön üye olarak homologdurlar. Aynı şekilde balina ve ayı balıklarının yassı üyeleri birbirinden bağımsız olarak fakat her ikisi de atasal bir karasal memelinin ön ayaklarından evrilmişlerdir. Dolayısıyla her ikisinin yassı üyeleri, diğer omurgalı ön üyelerindeki gibi aynı temel kemik yapısına sahip ön üyelerinin olması yönüyle homolog fakat yassı üyeleri modifikasyonlar homolog değil, anologdurlar.
Fenetik
Gördüğümüz gibi klasik taksonomistler hangi karakterlerin dikkate alınması gerektiği ve nasıl değer biçileceğine (ağırlıklandırma) ilişkin karar verirlerken kişisel yargıyı
kullanmalıdırlar. Sezgi bilimsel süreçte önemli rol oynamaktadır. Fakat klasik taksonomide subjektifliğin derecesi, birçok sistematikçiyi daha objektif bir yöntem geliştirmek için harekete geçirmiştir. Günümüzden 20 yıl kadar önce, daha az popüler olan yöntemlerden biri fenetiktir. Bu taksonomik yaklaşım, mümkün olduğunca fazla sayıda morfolojik karakter kullanır, tüm karakterlere aynı değeri biçer ve homoloji-anoloji karşıtlığı konusunu ihmal eder. Fosillerden embriyolojiden ve davranıştan edinilen kanıt sayısallaştırmak zor olduğundan genellikle hesaba katılmaz. Eğer yeterince karakter karşılaştırılırsa karakterlere oransal değer biçme ve anoloji durumlarının tanımlanması için gerekli olan subjektif yargıya gerek kalmayacağı beklenir. Diğer verilerin ağırlığı ile herhangi bir hata geçersiz hale gelecek ya da gizlenmiş olacaktır. Fenetikte (aynı şekilde burada tanımlanan diğer iki teknikte de), her bir karşılaştırmadaki farklılığın derecesi için bir değer oluşturularak türün her bir karakteri diğer türlerinkilerle karşılaştırılır. Daha sonra hesaplanan farklılık değerlerine karşılık gelecek şekilde, her bir tür, diğerlerine olan uzaklılıklarına göre yerleştirilerek dallanan bir diyagram oluşturulur. Böyle bir analiz, iki sorun nedeniyle karmaşıktır. İlki, böyle bir ağaç köksüzdür -yani hangi türün olası atasal form olduğu ya da evrimsel ağaçta dallanma noktalarının sırasına ilişkin gösterge yoktur. Her bir türün ortak atadan uzaklığını belirlemek için uygun bir grubun-analiz edilen gruba yakın akraba olan bir tür- hesaplamalara eklenmesi gerekir. İkincisi, hatalar belirsizlikler ve şans etkileri hesaplamalara ve hesaplamaya çalışılan evrimsel değişimlere karıştığından, farklı dal uzunlukları, hiçbir zaman bir dal çiftinden diğer dal çiftine (çataldan çatala) tamamen uymaz. Bazen bu kaçınılmaz anomaliler bazen çok küçük olması nedeniyle bir sorun oluşturmazlar, verilerle uyuşan yalnızca bir diyagram vardır ve değişik eşleştirmelerden çıkarılan uyuşmayan uzaklıkların basitçe ortalaması alınabilir. Fakat pek çok tür çalışılıyorsa ve dallanma noktaları birbirine çok yakın ise, belirsizliklerin olması olasıdır.
Teoride dal uzunlukları, sürüklenme ve seçilimin bir sonucu olarak birikmiş farklılıkları yansıtır. Eğer yalnızca sürüklenme işlemişse en dipteki dal noktasından, günümüz her bir türüne olan toplam uzaklık, en son ortak atadan beri geçirilmiş olunan zamanı yansı tarak aynı olacaktır. Sapmalar -yani daha uzun yollar alışılmadık seçici baskı derecesini ve grupta evrimleşmenin temposu ve modeli hakkında bir şeyler gösterir.
Fenetik (çoğunluk numerik taksonomi de denir) çoğunlukla sıkça oluştuğu gibi iki varsayımı -filogeni belirlemede bütün karakterler aynı derecede kullanışlı olduğu ve konvergent evrimleşmelerin çok az oluştuğu durumlarda ciddi problemlerle karşılaşılır. Örneğin; fenetik, harfi harfine uygulanırsa, plesantalı ve keseli yakın bir akraba olarak sınıflandırılır. Bu fenetikçilerin kendilerinin kabul edemiyecekleri bir sonuçtur. Daha sonra göreceğimiz gibi moleküler yaklaşımlar, fenetik matematiksel teknikleri, daha az bir hata riski ile kullanırlar.
Kıladistik Kıladistikçiler ortak türemiş karakterlere (çalışılmakta olunan grupta birçok tür için ortak olan ve eski ortak atalarından daha sonra oluşmuş olan özelliklere) yoğunlaşarak, anoloji ve homolojiyi karıştırmamaya özen gösterirler. Yani iki yarasa türünün sınıflandırılmasında, genelde memeliler tarafından paylaşılan karakterler dikkate alınmayacaktır, oysa fenetik, ölçülebilir tüm özellikleri değerlendirmeye tabi tutar. Kıladistik yaklaşım, bazı türlerin, kesin olarak dahil oldukları grupların genel taksonomik yapılarını bulundurmaması durumunu (örneğin sucul memelilerin arka üyelerinin yitirilmesi) göz ardı etmesi açısından bir kural getirdiği için yararlıdır. Kıladistikçi, değerlendirdiği karakterlerin herbirine, eşit oranda değerlendirilmeye alır, fakat incelenen türleşme olayının oluştuğu noktada paylaşıldığı var sayılan karakterleri gözardı ederek, fenetikçilerden çok daha seçici olabilirler: kullanılacak karakterlerin seçimi için kıladistikçilerin kural ya da ölçütleri vardır. Ancak hangi karakterlerin paylaşılan türemiş karakterler olduğu halen subjektif bir yargıyı çağrıştırdığından, bu yönü eleştirilebilir. Fenetik gibi kıladistik analiz genellikle fosilleri, embriyolojiyi ve davranış gözardı eder. Sonuç bazen tartışmalıdır. Örneğin; kıladistik analiz, timsahları kuşlara, memelileri ise yılan ve kaplumbağalara yakın gösterir. Bu görüş, gelişmiş omurgalıların doğru filogenisini yansıtsın ya da yansıtmasın, omurgalıların bu evrimsel versiyonunu biyologlar tarafından pek kabul görmemektedir. Fenetik ve klasik taksonomi gibi, kıladistik analiz de Avustralya keselilerinin gösterdiklerindeki gibi konvergenslerle yanıltılabilir. Bununla birlikte, yakın akraba cins ve türlerin filogenilerini analiz etmek için kıladistik yöntem yaygın olarak kullanılabilir.
Moleküler Taksonomi
Organizmaları sınıflandırmadaki en popüler yeni yöntem moleküler taksonomidir. Bu  yaklaşım, moleküler seviye üzerinde yoğunlaşarak homoloji/anoloji tartışmasından uzak durur. Moleküler konvergent evrimleşme olasılığının çok az olduğunu ve dolayısıyla bu seviyedeki analizlerin daha doğru olacağını varsaymaktadır. Ölçümler çeşitli şekillerde yapılabilir. Tekniklerden biri, iki türün DNA’sını tek zincir molekül halinde denatüre etmek, bunları birbirine karıştırmak ve çift zincirli melezleri oluşturmalarına izin vermektir. Hibridizasyon seviyesi ne kadar fazla ise iki türün o kadar yakın akraba oldukları varsayılır. En azından kuşlardan daha az çeşitlilik gösteren gruplarda bunun oldukça iyi çalıştığı görülmektedir. Bununla birlikte tekrarlanan DNA bölgelerinin hızlı evrimleşmesi, bu prosedürün ölçmeye çalıştığı sürüklenme ve seçilimden kaynaklanan farklılıkları etkileyebilir ve bu faktörü düzeltmek için kullanılan en iyi yöntemler üzerinde bile bazı tartışmalar vardır.
yaklaşım, moleküler seviye üzerinde yoğunlaşarak homoloji/anoloji tartışmasından uzak durur. Moleküler konvergent evrimleşme olasılığının çok az olduğunu ve dolayısıyla bu seviyedeki analizlerin daha doğru olacağını varsaymaktadır. Ölçümler çeşitli şekillerde yapılabilir. Tekniklerden biri, iki türün DNA’sını tek zincir molekül halinde denatüre etmek, bunları birbirine karıştırmak ve çift zincirli melezleri oluşturmalarına izin vermektir. Hibridizasyon seviyesi ne kadar fazla ise iki türün o kadar yakın akraba oldukları varsayılır. En azından kuşlardan daha az çeşitlilik gösteren gruplarda bunun oldukça iyi çalıştığı görülmektedir. Bununla birlikte tekrarlanan DNA bölgelerinin hızlı evrimleşmesi, bu prosedürün ölçmeye çalıştığı sürüklenme ve seçilimden kaynaklanan farklılıkları etkileyebilir ve bu faktörü düzeltmek için kullanılan en iyi yöntemler üzerinde bile bazı tartışmalar vardır.
Diğer bir teknik, proteinlerin amino asit dizisini karşılaştırır. Burada, çoğu yedek amino asitin nötr olması –örneğin nonpolar bir peptidin bir diğerinin yerini alması- ve dolayısıyla farklılıkların zamanla çok yavaş birikmesi gerekeceği varsayılır. Nerede kuvvetli seçilim baskıları varsa (örneğin dağlarda ya da derinlerde yaşayan sucul memelilerin hemoglobinleri üzerinde işleyen seçilim gibi), burada farklılıkların, sürüklenmeden beklenenden çok daha fazla olması olasıdır.
Son olarak, doğrudan DNA dizileri karşılaştırılabilir. Bu çok zahmetli bir iştir; fakat, rRNA genlerinin dizilerinin çıkartılması, alem (kingdom) ve şube (filum) seviyesindeki kategorilerle ilgili düşüncelerimizde tam bir devrim oluşturmuştur.
Her hangi bir proteini kodlayan genin kodonlarının üçüncü bazlarındaki değişimlerin çalışılması (aynı peptidin kodu olmaları nedeniyle çoğu nötr olan bu değişimler) cins, hatta tür içinde ayrıntılı analizlere olanak sağlar. Drosophila’nın filogenisi bu yöntem kullanılarak çalışılmıştır. Başlangıçta, gen dizisi analizlerinden ölçülen tüm şeylerdeki farklılıkların sayısının, dallanma noktalarının zamanlarını kesin olarak verecek bir moleküler saat olarak kullanılabileceği beklenmiştir. Ancak, aynı organizmadaki farklı genlerin çok farklı hızlarda değişecekleri, aynı şey farklı türlerdeki aynı genler için de geçerlidir. Şimdilik çok sayıda farklı genin ortalamasının işaret ettiği tarihler bile oldukça yaklaşık kabul edilmelidir.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç