
Bir astrositin hücre gövdesi, 10-20 mikronu kapsar ve işlemleri, bir 20-30 mikron boyunca yayılır ve stellat glial hücre oluşturur. Astrositinin bu yıldız şekli, ışık mikroskobu görüntüsünde daha net anlaşılabilir.
Astrositler, hipokampüsün CA1 bölgesindeki stratum radiatum hacminin % 4 ile 8’ini doldurur ve bu da onları en baskın glial alt tipi yapar. Astrositlere ek olarak; hipokampus, mikroglia ve oligodendrositler dahil olmak üzere 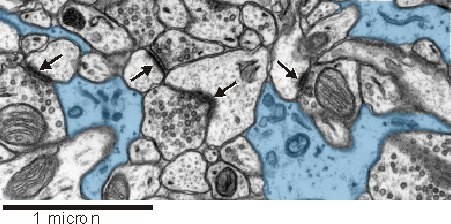 diğer glialara ev sahipliği yapar.
diğer glialara ev sahipliği yapar.
Sinapslara yakın astrositik bir varlık, sinaptik fonksiyon için önemli gibi görünmektedir (aşağıya bakınız). Sıçan hipokampüsünden elde edilen bu elektron mikrografında, sinaptik kompleksleri (oklar) ile yakından özdeşleşen astrositler (mavi) görülebilir.
Astrositler, en üst düzeyde yapısal özellikler ile tanımlanabilir:
• düzensiz, yıldız şeklinde
• sayısız glikojen granülü halinde
• ara filament demetleri olarak
• nispeten açık bir sitoplazma durumunda
Diğer glial alt tipleri gibi astrositler de, beyin işlevindeki asıl aktörler olan nöronlar için yalnızca destek ve bakım hücreleri olarak işlev gördüğü düşünülmektedir. Bununla birlikte geçtiğimiz birkaç yıl boyunca araştırmalar sinaptik olmayan ve aynı zamanda sinaptik iletişimdeki astrositler için çok daha merkezi bir rol olduğunu göstermiştirler.
Sinaptik iletimde astrositler, hacim, bileşim ve iyonların, nörotransmiterlerin, nöromodülatörlerin ve hücre dışı alandaki enerji kaynaklarının, konsantrasyonlarının modülasyonu yoluyla bir etki yapabilir. Mesaj iletimine astrositlerin dahil olması, muhtemelen sadece sinaptik öncesi ve sonrası ortakları değil, aynı zamanda komşu glia’yı da içerdiği gibi, sinaptik birimin yeniden gözden geçirilmesini sağlar.
Astrositik Sinsityum: Sinaptik Olmayan Bir İletişim Aracı
Sinaptik olmayan mesaj vericileri olarak astrositler, nöropilde astrositik sinsityum adı verilen bir ağ oluştururlar . Moleküllerin difüzyonu, sinsityumda astrositler arasındaki boşluk bağlantılarıyla meydana gelir.
Araştırmalar, kalsiyum dalgalarının sinsityum içinden yayıldığını (Cornell-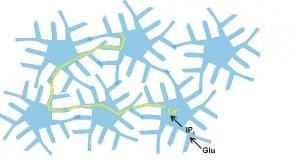 Bell ve diğerleri, 1990) ve bu dalgaların mekanik stimülasyon ve nörotransmitter glutamat tarafından indüklenebildiğini buldu. Kalsiyum dalgaları, ağ boyunca kalsiyum akışının astrositlere akması, kalsiyum duyarlı salınım ve iyonların ve nöromodülatörlerin alımına yol açarsa, terkibin uzun mesafe modülasyonuna ve hücre-dışı alandaki molekül konsantrasyonuna yol açabilir. Astrositik sinsityum, beyin içinde sinaptik olmayan bir iletişim aracı sağlar. Astrositler, birkaç sinapstan gelen bilgileri aynı anda algılayabilir ve entegre edebilir ve ayrıca hücre dışı alanın ve kan damarlarının içindeki kompozisyon hakkında bilgi alabilir. Astrositler, nöronların aksine, kuvvetlendirilemezler.
Bell ve diğerleri, 1990) ve bu dalgaların mekanik stimülasyon ve nörotransmitter glutamat tarafından indüklenebildiğini buldu. Kalsiyum dalgaları, ağ boyunca kalsiyum akışının astrositlere akması, kalsiyum duyarlı salınım ve iyonların ve nöromodülatörlerin alımına yol açarsa, terkibin uzun mesafe modülasyonuna ve hücre-dışı alandaki molekül konsantrasyonuna yol açabilir. Astrositik sinsityum, beyin içinde sinaptik olmayan bir iletişim aracı sağlar. Astrositler, birkaç sinapstan gelen bilgileri aynı anda algılayabilir ve entegre edebilir ve ayrıca hücre dışı alanın ve kan damarlarının içindeki kompozisyon hakkında bilgi alabilir. Astrositler, nöronların aksine, kuvvetlendirilemezler.
Nöronlar İçin Metabolik Faktörler Sağlanması
Astrositlerin sinaptik fonksiyona katıldığı bir yol, nöronal aktivite için metabolitlerin sağlanmasıdır. Beynin ana enerji kaynağı olan glukoz, merkezi sinir sistemine kan damarlarıyla temas eden astrositler yoluyla girer. Kan damarlarına temas eden astrositik süreçler astrositik son ayaklar olarak bilinir.
Kanıtlar, glukozun kısmen astrositler tarafından metabolize edildiğini ve ara maddelerin nöronal kullanım için serbest bırakıldığını gösterir 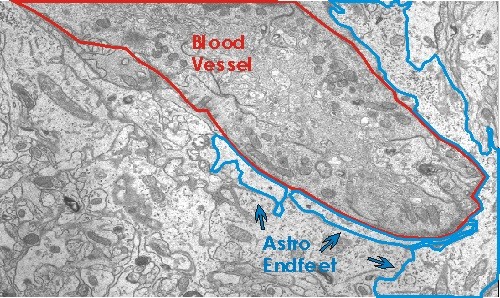 (Pfrieger ve Barres, 1996; Tsacopoulos ve Magistretti, 1996). Hipokampusta, glial metabolizma Krebs döngüsünün glia spesifik bir inhibitörü olan florositrat tarafından bloke edildiğinde, sinaptik geçiş engellenir (Keyser ve Pellmar, 1994; Pfrieger ve Barres, 1996). Bu, nöronların sinaptik işlev için gerekli olan yakıt için gliaya bağımlı olduğu anlamına gelir. Ek olarak, kanıtlar glia tarafından nöronlara verilen yakıt miktarının sinaptik aktivite miktarına bağlı olduğunu göstermektedir. Bir bulguya göre, glutamat astrositik glikoz alımını ve ardından laktatın hücre dışı boşluğa salınmasını uyarır (Pellerin ve Magistretti, 1994; Pfrieger and Barres, 1996).
(Pfrieger ve Barres, 1996; Tsacopoulos ve Magistretti, 1996). Hipokampusta, glial metabolizma Krebs döngüsünün glia spesifik bir inhibitörü olan florositrat tarafından bloke edildiğinde, sinaptik geçiş engellenir (Keyser ve Pellmar, 1994; Pfrieger ve Barres, 1996). Bu, nöronların sinaptik işlev için gerekli olan yakıt için gliaya bağımlı olduğu anlamına gelir. Ek olarak, kanıtlar glia tarafından nöronlara verilen yakıt miktarının sinaptik aktivite miktarına bağlı olduğunu göstermektedir. Bir bulguya göre, glutamat astrositik glikoz alımını ve ardından laktatın hücre dışı boşluğa salınmasını uyarır (Pellerin ve Magistretti, 1994; Pfrieger and Barres, 1996).
Astrositik Şişme ve Filopodia Uzatma ile Modüle ECS Hacim Fraksiyonu
Astrositler, hücre dışı boşluğun hacim fraksiyonunun önemli bir belirleyicisidir. Astrositik şişmenin hem glutamat hem de adenosin reseptörü stimülasyonunun bir sonucu olarak ortaya çıktığı gösterilmiştir (Hansson, 1994; Bourke ve ark. 1983). Filopodial genişleme veya membranın büyümesi astrositlerde glutamatın fokal uygulamasıyla indüklenebilir (Cornell-Bell ve diğerleri, 1992). Bu filopodial uzatma astrositik boyutunu ve morfolojisini değiştirebilir, muhtemelen hücre dışı boşluğun kapladığı hacim fraksiyonunu da değiştirebilir.
Sinaptik fonksiyon, hücre dışı hacim fraksiyonuna duyarlıdır. Hacim fraksiyonu azaldıkça hücre dışı boşluğun bileşenlerinin konsantrasyonu artar ve bu, teorik olarak, bir nöronun girilen belirli bir nörotransmitter miktarına tepkisini arttırır. Aslında, hücre dışı hacim fraksiyonunun astrositik şişmeye neden olduğu bilinen K+ konsantrasyonunun arttırılmasının bir sonucu olarak düştüğü zaman, nöronların uyarılabilirliğinin, nöbetlerin ortaya çıkma derecesine kadar arttığı 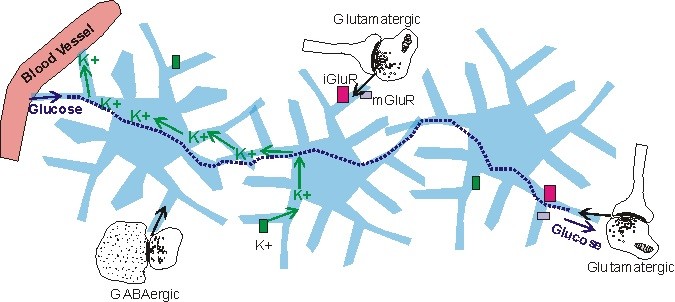 gösterilmiştir (Porter ve ark. McCarthy, 1997; Traynelis ve Dingledine, 1989). Ayrıca, astrositik şişmeyi engellediği bilinen furosemid’in yakın zamanda elektrikle indüklenen nöbetleri önlediği gösterilmiştir (Hochman ve diğerleri, 1995; Porter ve McCarthy, 1997).
gösterilmiştir (Porter ve ark. McCarthy, 1997; Traynelis ve Dingledine, 1989). Ayrıca, astrositik şişmeyi engellediği bilinen furosemid’in yakın zamanda elektrikle indüklenen nöbetleri önlediği gösterilmiştir (Hochman ve diğerleri, 1995; Porter ve McCarthy, 1997).
İyon Kanalları ve Nörotransmitter Reseptörleri
Astrositler, çok çeşitli iyon kanalları ve nörotransmitter reseptörleri ve taşıyıcıları içerir. K+ ve Ca2+ iyon kanalları bu iyonlar için beyin homeostazını korumak için astrositler tarafından kullanılır. Sinaptik aktivite sırasında, hücre dışı alanda K+ birikimi vardır, bu astrositik emilim ile azalır (Nilsson ve Hagberg, 1997). Astrositler daha sonra alınan K+’yı kılcal kanın içine fırlatır (Nilsson ve Hagberg, 1997). Astrositik Ca2+ iyon kanallarının rolü henüz net değildir.
İyon kanallarına ek olarak, astrositler nörotransmitter reseptörleri içerir. 1980’den önce (Van Calker ve Hamprecht, 1980), bu reseptörler sadece nöronlarda gözlendi ve astrositlerde olduğu düşünülmedi. Astrositlerde bulunan reseptör tipleri, nöronlarda bulunanlara benzer. Astrositik reseptörleri arasında, her biri astrositler arasında ve farklı beyin bölgelerinde farklı ekspresyon yapan ve her biri ikinci bir haberci kaskadıyla bağlanan (Kimelberg, 1995; gözden geçirilmiş; Porter ve McCarthy, 1997) glutamaterjik, gabaerjik, adrenerjik, serotonerjik ve muskarinik reseptörler bulunmaktadır.
Nöronal Uyarılabilirliği Etkileme Olasılıkları
Değişen hücre hacmine ek olarak, astrositlerin nöronal uyarılabilirliği modüle etmenin başka yolları da vardır. Örneğin, bir presinaptik terminale yakın bir astrositik depolarizasyon, salınan nörotransmitter miktarında bir artışa yol açabilir (Ronnback ve Hansson, 1997). Ayrıca, eğer verici salınımı Ca2+’ya bağımlıysa, hücre dışı alanda Ca2+ konsantrasyonunun astrositik kontrolü uyarılabilirlikte değişikliklere yol açacaktır (Ronnback ve Hansson, 1997). Başka olasılıklar da var. Aslında araştırma, kültürde gelişen nöronların glia ortaya çıkana kadar tam olarak işlevsel sinapslara ulaşmadığını göstermiştir (Pfrieger ve Barres, 1997), bunun için hareket mekanizması bilinmemektedir.
Astroglial Glutamat Alımı ve Sinaptik Glutamat Yayılımı Olasılığı
Astrositler, hücre dışı glutamat konsantrasyonunun sub-eksitotoksik seviyelerde tutulması ve böylece nöronal hücre ölümünün önlenmesi için kritik olan yüksek afiniteli glutamat taşıyıcıları içerir (Rothstein ve arkadaşları, 1994; Rothstein ve arkadaşları, 1996). Taşıyıcılar tarafından yetersiz glutamat alımının, amotrofik lateral skleroz, Alzheimer hastalığı, şizofreni ve AIDS’de rol aldığına inanılmaktadır. Asttamitik glutamat alımı, aynı zamanda, belki de sinaptik sinyali sonlandırmak suretiyle, sinaptik 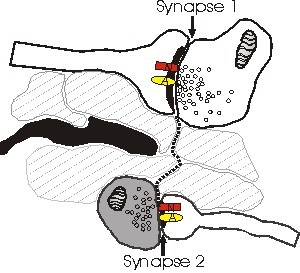 yarıkta glutamatın zaman sürecini ince ayarlamaya hizmet edebilir (Mennerick ve Zorumski, 1994). Ek olarak, astrositler, sintaztik glutamat dökülmesine aracılık edebilir.
yarıkta glutamatın zaman sürecini ince ayarlamaya hizmet edebilir (Mennerick ve Zorumski, 1994). Ek olarak, astrositler, sintaztik glutamat dökülmesine aracılık edebilir.
Yukarıda tasvir edilen, NMDA reseptörlerinin (N) neden AMPA (A) reseptörlerinden farklı bir kutata algıladıklarını açıklayan varsayımsal bir modeldir. Noktalı çizgi, sinaps 1’in yarığından kaçarken ve sinaps 2’ye ulaştığında glutamatın yolunu gösterir. Bu modelde, yayılma meydana gelir çünkü relase edilen glutamatın alınması yolu boyunca hiçbir astrositik işlem (siyah) yoktur. Sinaps 2’nin boğması (koyulaşmış) az miktarda veya hiç glutamat salmaz ve bu nedenle sinaps 2’ye ulaşan glutamat konsantrasyonu sadece (yüksek afiniteli) NMDA reseptörlerini aktif hale getirmek için yeterlidir. Bu model ile ilgili bir problem, yüksek seviyelerde ekstrasinaptik glutamatın genel bir eksitotoksisiteye yol açmasıdır.
Astrositler, glutamatın bunlardan yayılmasını etkili bir şekilde önlemek için yeterince yüksek yoğunlukta glutamat taşıyıcıları içerdiğinden, sinapsların yakınında astrositlerin varlığı, yayılmanın meydana gelip gelmediğinin önemli bir belirleyicisi olabilir. Hipokampal CA1 bölgesinde, glutamatı almak için herhangi bir araya giren yapı olmaksızın kısa sinaptik mesafeler gerçekten gözlenmiştir (Ventura ve Harris, 1999). Alternatif olarak, astrositler salınan glutatata doğru büyüdüklerinden (Cornell-Bell ve diğerleri, 1990), yalnızca akson-omurga arayüzlerinde astrositik süreçlerle sinapsların glutamat salgılaması mümkündür.
Yukarıda tasvir edilen NMDA reseptörlerinin (N) neden AMPA reseptörlerinden (A) farklı bir kutata algıladıklarını açıklayan alternatif bir modeldir. (A) ‘daki Sinaps 2, sintaz öncesi glutamat salgılayan az miktarda 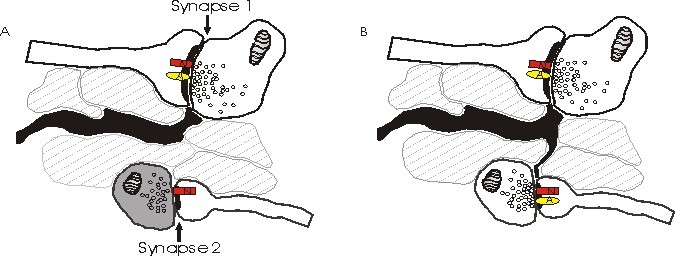 veya hiç yoktur ve sadece sindiyak olarak NMDA reseptörleri bulunur. Sinaps 2 kendiliğinden bir miktar glutamat salgıladığında, NMDA reseptörleri bunu algılar, ancak glutamatı algılayacak hiçbir AMPA reseptörü yoktur. (A) ‘daki sinaps 2’deki salıverme miktarı, komşu sinapsların üzerine dökülmek için yetersizdir ve astrositler (siyah), aktif sinaps 1’den dökülmesini önler. yeni çıkan sinaps 2’ye doğru. Eş zamanlı olarak sinaps 2, sinaps sonrası AMPA reseptörlerini işe aldı.
veya hiç yoktur ve sadece sindiyak olarak NMDA reseptörleri bulunur. Sinaps 2 kendiliğinden bir miktar glutamat salgıladığında, NMDA reseptörleri bunu algılar, ancak glutamatı algılayacak hiçbir AMPA reseptörü yoktur. (A) ‘daki sinaps 2’deki salıverme miktarı, komşu sinapsların üzerine dökülmek için yetersizdir ve astrositler (siyah), aktif sinaps 1’den dökülmesini önler. yeni çıkan sinaps 2’ye doğru. Eş zamanlı olarak sinaps 2, sinaps sonrası AMPA reseptörlerini işe aldı.
Bu model, astrositik büyüme/AMPA alıcı alım modeli , yayılmaya karşı savunuyor. Bunun yerine, NMDA ve AMPA reseptörleri tarafından algılanan kanta sayısının tutarsızlığının, sinaps aktif hale geldikçe sinaptik sonrası yoğunluklara AMPA reseptörlerinin alınmasıyla açıklanması mümkündür (Malenka ve Nicoll, 1995’te gözden geçirilmiştir).
Kaynakça:
Britannica
Yazar: Tuncay Bayraktar