
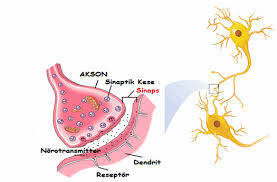
Klasik bir glutamaterjik sinaps, dendritik omurga ağacının en üstünde yer alır. Omurga, dendritik şafttan çıkıntı yapan bir boyun ve bilginin gerçekten alındığı bir kafadan oluşur. Glutamaterjik sinapsın genel bir açıklaması, postsinaptik omurgaya sahip bir yarıktan bakan bir presinaptik düğme içerir. Postsinaptik omurganın karşısındaki presinaptik düğmenin alanı, glutamatla dolu veziküller içerir ve aktif bölge (AZ) olarak adlandırılır. Salınmaya hazır vezikülleri yerleştiren bir protein kompleksi olan SNARE kompleksi (çözünür NSF bağlanma proteinleri) tarafından presinaptik membrana 10-20 arasında değişen bir dizi vezikül tutturulur.
Bir presinaptik sivri uçun gelişi, Ca2 + akışını takiben SNARE kompleksi tarafından aktive edilen vezikülün füzyonunu ve gözenek oluşumunu aktive eder. Tek temel bilgi bitinin transferinin ilk adımı, daha sonra bir presinaptik dikenin gelmesinin ardından SNARE kompleksi tarafından düzenlenen bir glutamat vezikülünün salınmasıdır. Spike’ı taşınan nöronal bilginin temel biti olarak düşünüldüğünde, EPSP’yi sinaptik olarak kodlanmış bilginin temel biti olarak düşünülebilir.
 Yerleştirilmiş vezikülleri içeren presinaptik yüzey, postsinaptik olandan 20 nm’lik bir mesafe (yarık) ile ayrılır. Sinaptik yarık, presinaptik vezikül tarafından salınan glutamat moleküllerinin Brown hareketi ile yayıldığı bir hacimdir. Ca 2+ ve SNARE kompleksi sayesinde presinaptik sivri uçların gelişi, bir vezikül ile presinaptik membran arasında bir gözenek oluşumuna neden olur. Bu gözenek, glutamat moleküllerinin vezikülden sinaptik yarığa geçerken izlediği yoldur.
Yerleştirilmiş vezikülleri içeren presinaptik yüzey, postsinaptik olandan 20 nm’lik bir mesafe (yarık) ile ayrılır. Sinaptik yarık, presinaptik vezikül tarafından salınan glutamat moleküllerinin Brown hareketi ile yayıldığı bir hacimdir. Ca 2+ ve SNARE kompleksi sayesinde presinaptik sivri uçların gelişi, bir vezikül ile presinaptik membran arasında bir gözenek oluşumuna neden olur. Bu gözenek, glutamat moleküllerinin vezikülden sinaptik yarığa geçerken izlediği yoldur.
Genel bir yatay kesit varsayarsak, kortikal veya hipokampal glutamaterjik sinapsın çapı 0.2-1 m arasında değişir. AZ dairesel uzay ve 20 nm’lik yarık varsayarsak, birçok yazarın bilgisayar modelleme yaklaşımı ile sinaptik iletimi incelemek için kullandığı silindirik bir alan hacmi elde edilir. Glutamatın difüzyonu için tüm sinaptik “silindir” serbest değildir. AZ, tüm sinapsın sadece bir kısmını kaplarken (ortalama yarıçap 0.11 m), çevreleyen kısım ise pre- ve postsinaptik nöronu sabitleyen fibriller tarafından işgal edilmiştir.
Postsinaptik tarafta, iki tip glutamat reseptörü, neredeyse AZ ile aynı boyutta olan ve dairesel bir şekil olarak kabul edilen bir alanda kolokalize edilir (silindirin alt kısmı). Bu alan postsinaptik yoğunluk (PSD) olarak adlandırılır ve -amino-3-hidroksi-5-metil-4-izoksazolpropiyonik aside duyarlı reseptörler (AMPA reseptörleri) ve siteleri içeren N-metil-d-aspartata duyarlı reseptörler (NMDA reseptörleri) içerir. glutamat moleküllerini bağlamak için (aynı zamanda sinaptik aktivite için gerekli olan glisin için). Bu iki tip reseptör, daha sonra detaylı olarak tartışacağımız sinaptik bilginin transferinde farklı rollere sahiptir. Her iki tip de dimerlerden oluşan tetramerlerdir ve reseptörün dimerik bileşimi postsinaptik tepkiyi şekillendirmede önemli bir rol oynar.
Görünüşe göre, bilgi aktarım süreci prensip olarak çok basittir. Presinaptik bir başağın gelişi, bir vezikülün, postsinaptik reseptörleri aktive eden ve dendritik dallardan yayılan EPSP adı verilen postsinaptik membran potansiyelinin bir varyasyonuna neden olan bir depolarize edici akım (EPSC) üreten glutamat salınımı ile füzyonunu üretir. (soma, postsinaptik başak oluşumuna) Bununla birlikte, presinaptikten postsinaptik tarafa geçişlerden herhangi biri, tüm süreci hem çalışmak hem de yorumlamak için son derece karmaşık hale getiren bir dizi bilgi yeniden düzenlenmesine uğrar.
Esasen, farklı modülasyon sistemleri postsinaptik yanıtın bir tür karmaşık doğrusal olmayan değişkenliğini üretir. EPSP’nin değişkenliğine hem pre- sinaptik hem de postsinaptik kontrol mekanizmaları neden olur ve bu değişkenliğin bir kısmı stokastik doğaya sahip gibi görünmektedir. Postsinaptik tepkinin değişkenliğinin ve bunların olası stokastik veya deterministik doğasının değiştirilmesini ima eden bazı (ancak hepsini değil) düzenleyici mekanizmalar vardır. Bu mekanizmalar şunlardır;
 EPSP değişkenliğinin ekstra sinaptik faktörleri
EPSP değişkenliğinin ekstra sinaptik faktörleri
Sınırlı sinaptik boşluğun hemen dışına bakıldığında, birkaç başka faktör EPSP oluşumunu etkileyebilir. Kısaca Denklemlere göre bunu söyleyenebilir. Sinapsın yakınlığındaki zar potansiyelini etkileyebilecek herhangi bir faktör EPSP’yi şekillendirmede rol oynayabilir. Sinaptik boşluğun dışında bulduğumuz ilk önemli yapı, omurganın boynu. Sinaps ve dendrit arasındaki iletişim yoludur ve elektrik direnci, hücreye (dendrit) geçen bilgi miktarını belirler. O halde boyun direncinin değeri, dendritlerin ve somanın farklı alanları arasındaki bilgi akışı için çok önemlidir. Omurga morfolojisi değişkendir ve dolayısıyla biyoelektrik özellikleri ve voltaj kapılı kanalların varlığı, sinaptik bilgiyi transfer etme yeteneğini daha da etkileyebilir.
Bazı araştırmacılara göre, boyun çapı ve direnci tek bir sinaptik olay sırasında da modüle edilir. Boyun direncinin modülasyonu, sonuç olarak, soma’ya iletilen EPSP’nin bir modülasyonunu üretir. Ancak boyun sadece sinaptik bilgiyi dendrite taşımaz. Dendrit durumu hakkındaki bilgileri PSD’ye taşıyarak da ters yönde hareket eder. Başka bir deyişle, PSD, yakınlarda bulunan diğer sinapslardan gelen bilgilerden haberdar edilir. Dendritik aktivite, aslında, dendrit ile omurganın başı arasında bir potansiyel farkı yaratarak, yönü iki yapı arasındaki potansiyel farkına bağlı olan net bir akım üretir. Dendritten gelen akım, esasen toplam EPSP’yi ve NMDA reseptörlerinin toplanmasını güçlü bir şekilde etkileyen PSD’nin yüksek giriş empedansı tarafından yükseltilir. O halde omurganın boynu, dendritlerdeki zar potansiyelinin uyarılma (depolarizasyon) seviyesine bağlı olarak sinaptik bilgi aktarımının güçlü bir modülatörüdür.
Çok sayıda girdiyi dikkate alarak ( 3×1033×103 -e 3×1043×104) alındığında, dendritik arborizasyon elektriksel bir izopotansiyel bölme değildir. Farklı branşlar arasındaki potansiyel farklılıkları, farklı girdi faaliyetlerinden ve meydana gelebildiği alanlarda geri yayılımın artmasından kaynaklanabilir. Spike geri yayılımı, dendritlerdeki Na + ve / veya Ca2 + voltaj kapılı kanalların varlığına bağlıdır. Bu kanalların varlığı ve yoğunluğu hem nöronlar arasında hem de aynı nöronda farklı dendritik bölgeler arasında farklılık gösterir. Ve sonuç olarak potansiyel farklılıkları dendritlerden geçen karmaşık potansiyel dalgalar üretebilir ve bu dalga, tek sinaptik olayı etkileyen boyun direnci yoluyla PSD’ye ulaşabilir.
 En son araştırmalarda, uyarıcı sinaptik aktivite tarafından üretilen potansiyel dalgaların, tekrar yayılmadan bağımsız olarak tek sinaptik yanıt üzerindeki olası bir etkisini incelenmiştir. Aktif sinaps sayısına ve bunların ortalama ateşleme frekansına bağlı olarak, tepkinin genliği, tepe seviyesi ve tepeye kadar geçen sürenin karmaşık doğrusal olmayan bir şekilde değiştiğini tespit edilmiştir. Aktif sinapsların sayısı, bir şekilde, örneğin, bir grup aktif nöronun, aynı nöron üzerinde aynı nöron üzerinde az çok senkronize bir şekilde (bir uyarana yanıt olarak) ateşlendiği beynin bir bölgesinden alınan girdiyi simüle eder. (dendritik alan)
En son araştırmalarda, uyarıcı sinaptik aktivite tarafından üretilen potansiyel dalgaların, tekrar yayılmadan bağımsız olarak tek sinaptik yanıt üzerindeki olası bir etkisini incelenmiştir. Aktif sinaps sayısına ve bunların ortalama ateşleme frekansına bağlı olarak, tepkinin genliği, tepe seviyesi ve tepeye kadar geçen sürenin karmaşık doğrusal olmayan bir şekilde değiştiğini tespit edilmiştir. Aktif sinapsların sayısı, bir şekilde, örneğin, bir grup aktif nöronun, aynı nöron üzerinde aynı nöron üzerinde az çok senkronize bir şekilde (bir uyarana yanıt olarak) ateşlendiği beynin bir bölgesinden alınan girdiyi simüle eder. (dendritik alan)
Daha önce bahsedilen durum için, örneğin, bir hipokampal alt sahanın bir nöronu, dentat girusun geniş bir alanından (birçok nöron) ve ayrıca dendritik dalların ayrı bölgelerindeki Entorhinal korteksin alanlarından senkronize girdiler alabilir. Bu iki alandan birinden gelen nöronlar, senkronizasyonlarının derecesine bağlı olarak ortalama bir frekans ve standart sapma ile ateşlenir. Böyle bir durum, aynı zaman aralığında aktif olan herhangi bir tek sinapsı doğrudan etkileyen uyarımla ilgilenen dendritik alana dalgalar üretir.
Alıcı nöronun membran potansiyeli, bir voltaj “bandı” oluşturan iki seviye arasında salınır. Bu voltaj “bandının” genliği, aktif sinapsların sayısına ve bunların ortalama ateşleme frekansına bağlıdır. Belirli bir sinapsın EPSP’si, “bant” içinde herhangi bir seviyede meydana gelebilir. Denklemlere göre, belirli bir sinapsın EPSP’sinin başladığı seviyeye bağlı olarak, özellikleri (genlik, tepe seviyesi, NMDA katkısı, vb.) değişecektir. Bu bantta, maksimum olasılık seviyesi olarak kabul edilebilecek bir ortalama değer belirlemek mümkündür.VmVmEPSP’nin meydana gelebileceği.
Bu ortalama seviye, aktif sinapsların sayısını veya ateşleme frekanslarını artırarak artar (daha depolarize) Bu voltaj “bandının” varlığı, olası seviyelerde büyük bir gama sağlar. VmVmEPSP’nin meydana gelebileceği ve sonuç olarak meydana gelme zamanına (bant içindeki salınımın fazı) bağlı olarak tek EPSP’nin çok güçlü bir düzenleyicisini temsil eder. Farklı bir şekilde söylenirse, EPSP’nin belirli bir seviye ile çakışması, sinapsın aktardığı bilginin türünü ve miktarını belirler. Aktiviteleri için membran voltajına bağlı olan NMDA reseptörleri özellikle bu tür bir düzenlemeye duyarlıdır ve aslında EPSP’nin diğer sinapsların aktivitesiyle “çakışması”, LTP ve hafıza gibi fenomenler için çok önemli kabul edilir.
(NMDA’ya bağlıdır) Bunlar, nöronların, çoğunlukla LTP ve hafıza fenomenlerini üretmede tesadüf dedektörleri olarak hareket ettiğini öne süren temel mekanizmalardırTek bir bit sinaptik bilginin aktarımı üzerindeki dendritik aktivite düzenleyici etki, esas olarak zardaki potansiyel varyasyonuna bağlıdır ve şu şekilde özetlenebilir: Artış geri yayılımı ve Dendritik ağaçta aktif sinaptik girdiler.
Kaynakça:
https://www.cell.com/fulltext/S0896-6273(03)00608-1
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4743082/
Yazar: Özlem Güvenç Ağaoğlu