
 DNA’nın eksiksiz bir biçimde kendini yenilemesi normal bir hücrenin işlevi için zorunludur. Mutasyonlar yani genlerde meydana gelen kalıtsal değişiklikler enzim yapısında değişime yol açarak metabolik bir yolu geliştirmeyip onu bozarlar. Yapısal protein, enzimler, düzenleyici proteinler gibi binlercesinin sentezi için gerekli olan gen komutları yüzlerce baz uzunluğunda olduğundan, baz başına binde birlik hata oranı önemsiz gibi görünse de aslında çok daha büyük bir orandır. Bu durumda hem prokaryot hem de ökaryotlarda bulunan mutasyonları tespit edip, onaran özel enzimlerin olması şaşırtıcı değildir.
DNA’nın eksiksiz bir biçimde kendini yenilemesi normal bir hücrenin işlevi için zorunludur. Mutasyonlar yani genlerde meydana gelen kalıtsal değişiklikler enzim yapısında değişime yol açarak metabolik bir yolu geliştirmeyip onu bozarlar. Yapısal protein, enzimler, düzenleyici proteinler gibi binlercesinin sentezi için gerekli olan gen komutları yüzlerce baz uzunluğunda olduğundan, baz başına binde birlik hata oranı önemsiz gibi görünse de aslında çok daha büyük bir orandır. Bu durumda hem prokaryot hem de ökaryotlarda bulunan mutasyonları tespit edip, onaran özel enzimlerin olması şaşırtıcı değildir.
Replikasyon Süresince Onarım
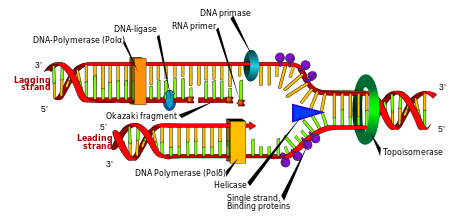 Hem prokaryotların hem de ökaryotlann replikasyon enzimlerinin DNA sentezindeki hata oranı 105 baz çifti için yaklaşık olarak 3’tür (Bu değer, bir ya da birden fazla alt üniteden meydana gelen kompleks yapıdaki DNA polimerazlarla ilgili çalışmalardan elde edilmiştir). Kompleks yapıdaki DNA polimeraz enzimlerinin yapılarından sadece hata tamirinden sorumlu enzim alt üniteleri çı karılarak ya da mutasyonlara bağlı olarak bu alt üniteleri çalışmayan DNA polimeraz enzimleri kullanılarak gerçekleştirilen DNA replikasyonlarında DNA sentezinin meydana geldiği; fakat atasal DNA dan farklı olarak birçok hatalı baz çiftinin oluştuğu saptanmıştır. Eğer yüksek oranda gerçekleşen bu hatalar düzeltilmeden bırakılırsa sonuç olarak her hücrenin proteinlerinde yaklaşık yüzde 3’lük bir hata olacaktır (belki de her insan hücresinde her replikasyon sonrası oluşmuş 1000 farklı ve değişik yapıda protein). Neyse ki DNA polimeraz kompleksi, sentezlenen her bazı tekrar tekrar gözden geçiren ve hatalı olanları çıkaran bir ya da daha fazla enzim alt ünitelerini içerir. DNA polimeraz kompleksindeki bu alt üniteler yanlış eşleşmiş baz çiftlerini tanır, yerine doğru eşleşen bazı koyarlar.
Hem prokaryotların hem de ökaryotlann replikasyon enzimlerinin DNA sentezindeki hata oranı 105 baz çifti için yaklaşık olarak 3’tür (Bu değer, bir ya da birden fazla alt üniteden meydana gelen kompleks yapıdaki DNA polimerazlarla ilgili çalışmalardan elde edilmiştir). Kompleks yapıdaki DNA polimeraz enzimlerinin yapılarından sadece hata tamirinden sorumlu enzim alt üniteleri çı karılarak ya da mutasyonlara bağlı olarak bu alt üniteleri çalışmayan DNA polimeraz enzimleri kullanılarak gerçekleştirilen DNA replikasyonlarında DNA sentezinin meydana geldiği; fakat atasal DNA dan farklı olarak birçok hatalı baz çiftinin oluştuğu saptanmıştır. Eğer yüksek oranda gerçekleşen bu hatalar düzeltilmeden bırakılırsa sonuç olarak her hücrenin proteinlerinde yaklaşık yüzde 3’lük bir hata olacaktır (belki de her insan hücresinde her replikasyon sonrası oluşmuş 1000 farklı ve değişik yapıda protein). Neyse ki DNA polimeraz kompleksi, sentezlenen her bazı tekrar tekrar gözden geçiren ve hatalı olanları çıkaran bir ya da daha fazla enzim alt ünitelerini içerir. DNA polimeraz kompleksindeki bu alt üniteler yanlış eşleşmiş baz çiftlerini tanır, yerine doğru eşleşen bazı koyarlar.
Kompleks hata tamirinden hemen sonra ikinci bir kez aynı bölgeyi tarayarak tamirin doğruluğunuda kontrol etmektedir. Bu ikinci denetleme hata oranını yaklaşık 109 da bire düşürür. Bu durum insanda DNA daki iki zincirin herbiri üzerinde ortalama olarak 1500 baz içeren 50.000 işlevsel genden bir tanesi her on hücre bölünme sonucunda hatalı olur. Görüleceği gibi bu oran genelde diğer mutasyon kaynaklarından daha küçüktür.
Diğer Tipdeki Mutasyonların Onarımı
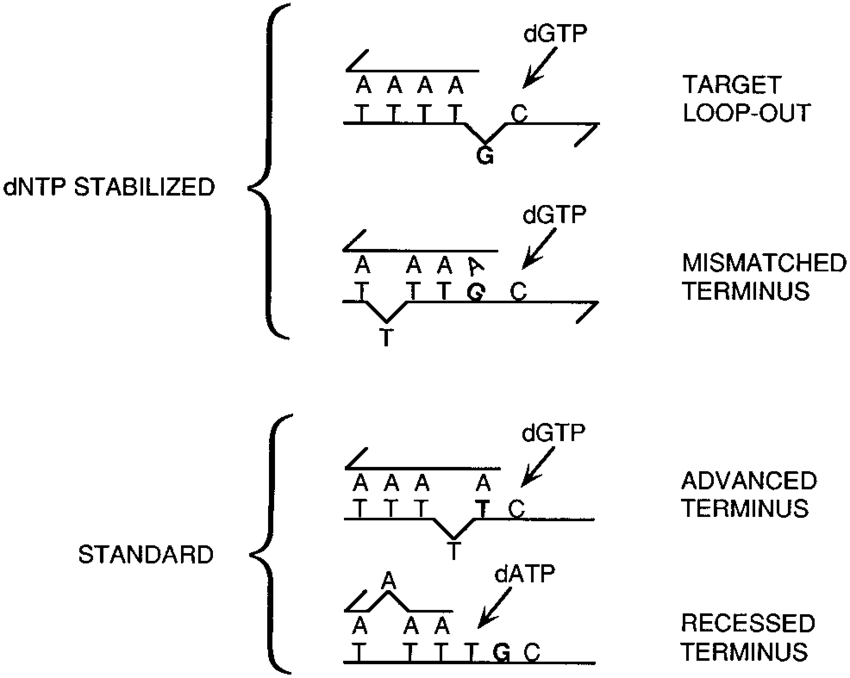 Genetik mesajın bütünlüğü aynı zamanda baz sekanslarında sıcaklık, radyasyon ve çeşitli kimyasal ajanların etkisiyle meydana gelen değişiklikler ile de tehdit altındadır. Bu mutasyonların oluşma oranı şaşılacak ölçüde yüksektir. Mesela, yalnızca termal enerji, her gün, her bir insan hücresindeki yaklaşık 5000 pürin (adenin-guanin) ile deoksiriboz iskeleti arasındaki bağı kırar. Sitozin, her gün kimyasal olarak bir hücrede 100 kez urasile (normalde sadece RNA’da bulunan ve DNA replikasyon ve transkripsiyon enzimlerince yanlış olarak okunan nükleotit) çevrilir.
Genetik mesajın bütünlüğü aynı zamanda baz sekanslarında sıcaklık, radyasyon ve çeşitli kimyasal ajanların etkisiyle meydana gelen değişiklikler ile de tehdit altındadır. Bu mutasyonların oluşma oranı şaşılacak ölçüde yüksektir. Mesela, yalnızca termal enerji, her gün, her bir insan hücresindeki yaklaşık 5000 pürin (adenin-guanin) ile deoksiriboz iskeleti arasındaki bağı kırar. Sitozin, her gün kimyasal olarak bir hücrede 100 kez urasile (normalde sadece RNA’da bulunan ve DNA replikasyon ve transkripsiyon enzimlerince yanlış olarak okunan nükleotit) çevrilir.
Güneş ışınlarının ultraviyole radyasyonu etkiye açık epidermal hücrelerde yüksek oranda aynı zincir üzerindeki komşu timinleri birleştirir. Onarıcı enzimlerin devamlı işlevlerinden dolayı mutasyonların hücrede ortalama toplanma sı klığı replikasyondaki düzeltilememiş hata oranından daha düşüktür.
Replikasyon hatalarında olduğu gibi birçok çeşit mutasyonda da kromozomlann onarılma stratejisi esas olarak aynıdır. Enzimler hatalı dizilimi tespit edip buraya bağlanırlar ve hatalı bazları yapıdan çıkarırlar. Bu sistemde atasal zincirlerden bozulmamış olan komplementer (tamamlayıcı eş) yapı zincir onarımını yönlendirir. Bir seri dikkat çekici özgül enzimler bu işlemde görev alır, örneğin, kromozomlar herbiri belirli sınıf problemlere özgü 20 değişik enzimle kimyasal baz değişimleri açısından taranır. 5 ila 10 civarındaki diğer bazı özel enzimler, bazlarla diğer kimyasal maddeler ya da aynı zincirdeki komşu bazlar arasındaki yanlış kurulmuş kovalent bağları tanıyıp tamir etmek için özelleşmiştir. Ultraviyole radyasyonu ile timinlerin birleşmesi bu tip hataların en çok rastlananıdır. Diğer bir grup enzim baz kaybı olan bölgelere (mesela pürinler termal enerjiyle kolaylıkla DNA yapısından koparak ayrılırlar) bağlanarak DNA tamirine katılırlar.
Bu özenli onarım sistemine karşın bazı mutasyonlar, mavi gözden (bozuk bir pigment geninin varlığı sonucu oluşur) kalıtsal genetik hastalıklara kadar olağanüstü değişimlere neden olacak şekilde ayakta kalmayı başarır. Bu nasıl gerçekleşir? Büyük bir olasılıkla enzimatik sistemlerin hiçbiri mükemmel değildir. Bazı hatalar gözden kaçar ve bazıları yanlış onarılır. Bunun yanında bir mutasyon replikasyon esnasında ya da daha sonra gerçekleştiğinde, bunu onarmak için yeterli zaman olmayabilir. Hala, diğer mutasyonlar tam olarak incelenmemiştir; fakat büyük bir olasılıkla onarım sistemince meydana getirilirler.
 Onarım enzimlerince meydana getirilen mutasyonların iyi tanınanlarından birisi misalignment delesyondur. Son çalışmalar göstermiştir ki, küçük delesyonlar (birkaç baz çifti kaybı ) gelişigüzel bölgelerde görülmemektedir; bazı baz dilimlerinin yeraldığı bölgeler delesyona diğerlerinden daha açıktır. Bu farklı duyarlılığın nedeni de, adenin ile timin baz çifti arasındaki oransal zayıflıktır. Sitozin ve guanin üç hidrojen bağıyla bağlıyken, onlar sadece iki hidrojen bağı ile bağlıdır. Hidrojenin bağları sürekli kırılıp tekrar yapılır ve bu şans eseri küçük bir DNA segmentinde tüm bağların aynı anda yıkılmasına neden olabilecek şekilde adenin timin baz çiftince zengin bölgelerde sıkça tekrarlanır. Bağlar aynı anda tekrar kurulurlar; fakat bazen tekrar bağlanma hatalıdır. Mesela, adenin timin bazlarını uzunca bir seri olarak içeren bir bölgede bu tip bir geçici bölünme olduğunda, iki zincir tekrar birleşirken, küçük bir ihtimalle de olsa yanlış çizgilenme gerçekleşebilir. DNA onarım enzimleri de eşleşmemiş bazları uzaklaştırıp sonuçta misalignment delesyona neden olabilir. Onarım bu durumda genin mRNA’sının translasyonu esnasında hataya neden olurken, genin “anlamını” değiştirmektedir.
Onarım enzimlerince meydana getirilen mutasyonların iyi tanınanlarından birisi misalignment delesyondur. Son çalışmalar göstermiştir ki, küçük delesyonlar (birkaç baz çifti kaybı ) gelişigüzel bölgelerde görülmemektedir; bazı baz dilimlerinin yeraldığı bölgeler delesyona diğerlerinden daha açıktır. Bu farklı duyarlılığın nedeni de, adenin ile timin baz çifti arasındaki oransal zayıflıktır. Sitozin ve guanin üç hidrojen bağıyla bağlıyken, onlar sadece iki hidrojen bağı ile bağlıdır. Hidrojenin bağları sürekli kırılıp tekrar yapılır ve bu şans eseri küçük bir DNA segmentinde tüm bağların aynı anda yıkılmasına neden olabilecek şekilde adenin timin baz çiftince zengin bölgelerde sıkça tekrarlanır. Bağlar aynı anda tekrar kurulurlar; fakat bazen tekrar bağlanma hatalıdır. Mesela, adenin timin bazlarını uzunca bir seri olarak içeren bir bölgede bu tip bir geçici bölünme olduğunda, iki zincir tekrar birleşirken, küçük bir ihtimalle de olsa yanlış çizgilenme gerçekleşebilir. DNA onarım enzimleri de eşleşmemiş bazları uzaklaştırıp sonuçta misalignment delesyona neden olabilir. Onarım bu durumda genin mRNA’sının translasyonu esnasında hataya neden olurken, genin “anlamını” değiştirmektedir.
DNA yapısındaki bazların metilasyonu gen ifadesinin düzenlenmesinde kullanılan yöntemlerden birisidir. Genomdaki bazı sitozinlerin metillenmesi olumlu bir uyum olabilir. Fakat bu olgu bazı durumlarda birtakım problemleri de birlikte getirir. Bakterilerde kısıtlayıcı endonükleazlar (restriction endonuclease) olarak bilinen sitoplazmadaki bazı enzimler, istilacı virüslerin DNA’larını parçalayarak bakterilerin savunma silahı olarak görev yapmaktadırlar. Bu tip enzimler istilacı DNA’yı parçalıyorsa, neden aynı zamanda bakteriyel kromozomu da kesmemektedirler?. Cevap ise bakteriyel DNA’da kısıtlayıcı endonükleazların tanıyıp kestiği baz dizilerindeki sitozinlerin sadece bu bölgelerde özgül olarak metillenmiş olmaları ve böylece bakterilerin kendi kısıtlayıcı endonükleazlannın aktivitelerinden korunmuş olmalarıdır. Fakat endonükleazların aktivitelerinden bu şekilde korunan bakteriyel DNA daki sitozinin amino grubunu yitirerek timine dönüşmesi durumunda mutasyon oluşması kaçınılmazdır. Okaryotlarda da, aktif olarak bazların metillenmesi söz konusudur ve işlevi gen regülasyonuna yardım etmektir.
Kaynakça:
Khan Academy
Yazar: Taner Tunç