
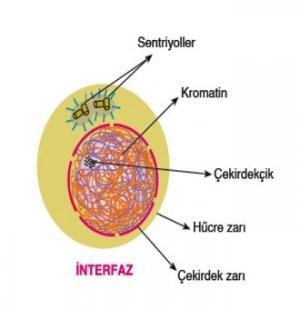
 İnterfaz mitoz ve mayoz bölünmenin ilk evresidir. Bölünmeyen hücrenin interfaz durumunda olduğu söylenir. Çekirdek, zarla çevrili bir organel biçiminde belirgin olarak görünmektedir ve bir ya da daha fazla çekirdekçik de kolayca görünebilir. Ancak kromozomlar, alışılmış biçimleriyle çekirdek içerisinde görünür değildirler; mikroskoplar ve ayrıntılı boyama tekniklerinin bölünen bir hücrede görmemizi sağladığı çubuk biçimli cisimlerden hiç biri ortada yoktur. İnterfaz kromozomları o kadar ince ve birbirinden ayırt edilmeleri öylesine olanaksızdır ki, yalnızca düzensiz bir kromatin yığınına benzemektedirler.
İnterfaz mitoz ve mayoz bölünmenin ilk evresidir. Bölünmeyen hücrenin interfaz durumunda olduğu söylenir. Çekirdek, zarla çevrili bir organel biçiminde belirgin olarak görünmektedir ve bir ya da daha fazla çekirdekçik de kolayca görünebilir. Ancak kromozomlar, alışılmış biçimleriyle çekirdek içerisinde görünür değildirler; mikroskoplar ve ayrıntılı boyama tekniklerinin bölünen bir hücrede görmemizi sağladığı çubuk biçimli cisimlerden hiç biri ortada yoktur. İnterfaz kromozomları o kadar ince ve birbirinden ayırt edilmeleri öylesine olanaksızdır ki, yalnızca düzensiz bir kromatin yığınına benzemektedirler.
İnterfaz geçiren hayvan hücrelerinde, çekirdeğin tam dışında yeralan ve birbirine dik açılı konumda iki küçük silindir biçimli cisim içeren özel bir sitoplazma bölgesi vardır. Bunlar sentriollerdir, ileride kendilerini eşleyecek, biri diğerinden ayrılacak ve bölünmekte olan hücrenin kutupları ile ilişkili hale geçeceklerdir. Kamçı ve siller bunlardan türediği için, sentriyollerin oluşacak kardeş hücrelere en az kromozomlar kadar güvenli ve hatasız bir biçimde aktarılması gerekir. Pek çok hayvan hücresinde, sentriyoller mitozdan hemen önce birbirinden ayrılır ancak bazı hücrelerde mitozun başlamasından oldukça önce, interfaz sırasında bu olay gerçekleşir. Tohumlu bitkilerin çoğunda sentriyole henüz rastlanılamamıştır. Bu durum pek şaşırtıcı değildir; çünkü bu türlerde kamçı ya da sil bulunmamaktadır. Ancak hareketli sperm üretilmesi ile bağlantılı olarak bazı alglerde, mantarlarda, biriyofitlerde ve eğreltilerde sentriyoller vardır. Belirli bir işlevi oldukları izlenimi verecek netlikle görünür hale geçmelerine karşın sentriyoller, hayvan hücrelerinin bölünmesi için bile yüzde yüz gerekli değildir; mitoz boyunca iğ iplikçiği düzenleyici merkezlerin yakınındadırlar; ama iplikçik oluşumunda rolleri yoktur.
![İnterfaz Evresi Nedir? İnterfaz Evresinde Gerçekleşen Olaylar]() İnterfaz Hücreleri
İnterfaz Hücreleri
 İnterfaz Hücreleri
İnterfaz HücreleriEskiden interfaz hücrelerine dinlenen (durgun) hücreler deniyordu; fakat şimdi bu yaklaşımdan uygun olmadığı için vazgeçilmiştir. Bir interfaz hücresi kesinlikle dinlenme durumunda değildir; işlev gösteren canlı bir hücrenin sahip olduğu sayısız etkinliğin hepsini gerçekleştirmektedir. Solunum, protein sentezi, büyüme, farklılaşma ve daha niceleri. İnterfaz sırasında, dahası, genetik madde çoğaltılmaktadır (replike edilmektedir) ve hücredeki tüm sistem ve sonraki bölünme evresine hazırlık olarak iki katına çıkarılmaktadır. Genetik maddenin replikasyonu (çoğaltılması ), bununla birlikte, son bölünme evresinin tamamlanmasının hemen ardından başlamaz.
Genetik replikasyondan önce G, evresi denen bir zaman aralığı vardır. Ribozomlar ve organeller iki katına çıkar. Ardından, organellerin iki katına çıkarılma işleminin de sürdüğü ve yeni DNA sentizinin gerçekleştiği S evresi gelir. G2 denen bir başka evre replikasyon bitimini mitoz başlangıcından tamamen ayırır. Bu zaman süresince hücre mitoza hazırlanmaktadır. İnterfazın bu üç alt evresi, mitoz ve sitokinez ile birlikte (ikisi birlikte M fazı olarak adlandırılır), hücre döngüsü’nü oluşturur. Doku kültüründeki hücreler hücre döngüsünden geçerken çarpıcı değişimler gösterirler, ancak bu değişimlerin, özellikle, bütünlüğünü koruyan bir organizma açısından neye işaret ettiği iyice bilinmemektedir.
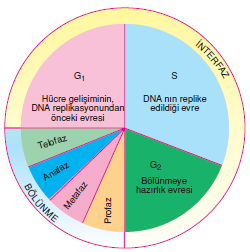 Bütün bu hücre döngüsünün süresi büyük farklılık gösterebilir. Bitkilerde geçen süre 10-30 saat ve hayvanlarda 18-24 saat olmasına karşın, bazı organizmalarda 20 dakika gibi kısa ya da günler hatta haftalar kadar uzun olabilmektedir. Bir dereceye kadar tüm evrelerin süreleri birbirinden farklıdır; ama en büyük farklılık G, evresinde olmaktadır.
Bütün bu hücre döngüsünün süresi büyük farklılık gösterebilir. Bitkilerde geçen süre 10-30 saat ve hayvanlarda 18-24 saat olmasına karşın, bazı organizmalarda 20 dakika gibi kısa ya da günler hatta haftalar kadar uzun olabilmektedir. Bir dereceye kadar tüm evrelerin süreleri birbirinden farklıdır; ama en büyük farklılık G, evresinde olmaktadır.
Bir taraftan son derece hızlı bölünen ve G, evresi yoktur denebilecek çabuklukla bu evreyi geçebilen embriyo hücreleri gibi hücreler varken, öte yandan G, evresinde tutulu kalan bazı hücre tipleri söz konusudur. Örneğin farklılaşmalarını tamamlanmış iskelet kası hücreleri ve sinir hücreleri G, evresinde kalmışlardır ve normalde asla yeniden bölünmezler. Hücrelerin G2 evresinde tutulu kaldıkları bir kaç durum vardır; hücreler DNA’larını replike etmişlerdir; ama bölünmezler. G2 de tutulu kalan hücrelere bir örnek yetişkin insanın kalp kas hücreleridir. G2’de tutulu kalma olgusu günümüzde net bir açıklamaya sahip değildir. G1 ve G2 de tutulu kalışın ortak nedeni temel bir kontrol maddesinin üretimindeki bir hatadır. Glevresinde kalmış bir hücrenin çekirdeği S fazına girmekte olan bir hücreye aktarıldığında, aktarılan bu çekirdek hemen etkinleşecek ve kendisi de S fazına girecektir. Çünkü girdiği hücrenin sitoplazmasında bulunan bir kontrol maddesi tarafından uyarılmıştır. Benzer biçimde, G2 de kalmış bir hücre bir mitoz hücresi ile kaynaştırılırsa kromozomları hemen yoğunlaşmaya başlar ve kendisi de mitoza girer.
Siklin
 Hücre döngüsü kontrol eden maddelere siklin denir. S-siklin replikasyonu (DNA sentezi) uyarır, M- siklin mitozun başlamasına yardım eder. Her birinin bağlandığı hücre bölünmesi döngüsü proteini (cdc) aynıdır. Bu protein hücredeki, replikasyon ya da mitoza özgü bir mesajcıyı etkinleştirecektir.
Hücre döngüsü kontrol eden maddelere siklin denir. S-siklin replikasyonu (DNA sentezi) uyarır, M- siklin mitozun başlamasına yardım eder. Her birinin bağlandığı hücre bölünmesi döngüsü proteini (cdc) aynıdır. Bu protein hücredeki, replikasyon ya da mitoza özgü bir mesajcıyı etkinleştirecektir.
Sistemin nasıl işlediğini görmek için tüm döngüyü G, fazıyla başlayarak takip edelim. G1 başlarken cdc proteini etkisiz, cdci, formundadır. Eğer hücre tekrar bölünecekse, S- siklin rutin biçimde sentezlenir. Üretim ne kadar çabuk olursa o denli hızlı bir biçimde S fazı başlayacaktır. S- siklin konsantrasyonu yükseldikçe cdci’ye bağlanmaya başlar ve onu sentez hızlandırıcı form olan cdcs’ye dönüştürür. Bir eşik durumuna ulaşıldığı zaman cdc etkinleşir ve bir mesajcı bileşiğe fosfor takmaya başlar. Bu şekilde etkinleşen mesajcı replikasyonun başlamasına yol açar ve öbür enzimler hemen S- siklini parçalar, cdcs etkisiz formu olan cdci formuna tekrar dönüşür.
G2 fazında, mitoz başlamasına yol açıcı M-siklin rutin biçimde sentezlenir. M siklinin konsantrasyonu yükseldikçe cdci’ye bağlanır ve onu mitoz hızlandırıcı cdcm formuna dönüştürür. Yıne, bir eşik durumundan geçildiğinde cdcm etkinleştirilir ve farklı bir mesajcı mitozun başlamasına ve M-siklinin parçalanmasına neden olur, cdcm cdci’ye dönüşür.
Kaynakça:
Khan Academy
Yazar: Taner Tunç