
 Yeni oluşmuş bir sinir hücresinin yaşamındaki ilk adım, gelişen bir organizmadaki diğer bir çok hücrelerdeki gibi, genellikle meydana geldiği yerden, bulunması beklenilen yere harekettir. Örneğin, retinayı oluşturacak hücreler, gelişmekte olan beyinden çıkarak gözlerin yerleşeceği yere doğru ilerleyip, daha sonra optik keseciği oluştururlar. Halbuki, beyinin dış tabakasındaki serebral korteks hücreleri yaşlı hücre tabakaları arasından oluştukları yer olan beyinin merkezinden, ait oldukları korteksin dış tabakasına ulaşmak için hareket etmelidir.
Yeni oluşmuş bir sinir hücresinin yaşamındaki ilk adım, gelişen bir organizmadaki diğer bir çok hücrelerdeki gibi, genellikle meydana geldiği yerden, bulunması beklenilen yere harekettir. Örneğin, retinayı oluşturacak hücreler, gelişmekte olan beyinden çıkarak gözlerin yerleşeceği yere doğru ilerleyip, daha sonra optik keseciği oluştururlar. Halbuki, beyinin dış tabakasındaki serebral korteks hücreleri yaşlı hücre tabakaları arasından oluştukları yer olan beyinin merkezinden, ait oldukları korteksin dış tabakasına ulaşmak için hareket etmelidir.
Gelişme sırasında sinir hücrelerinin hareketi iyi düzenlenmiştir, rastgele hareket etmezler. Böbreğin adrenal bezlerini ve sinir hücre yığınlarını (gangliyonları) oluşturacak hücreleri düşünün. Bu hücreler omuriliğin hemen üstünde bulunan nöral krestten kök alıp, aşağıya doğru notokordun hemen yanındaki özel bölgelere göç ederler. Oluştukları yerden uygun yöne doğru hareket edip, önceden belirlenmiş yolları izleyerek belirli bir noktada dururlar. Benzer bir şekilde, serebral korteks (beyin kabuğu) hücreleri dokunun bazal tabakasında meydana gelir ve sonra korteksin dış tabakasına ulaşana dek daha yaşlı kortikal hücrelerin arasından dışarıya doğru göç ederler. Nöral krest ve kortikal hücrelerin bir yerden başka bir yere geçişindeki üç evre-başlangıç yönünün belirlenmesi, bir yolun izlenmesi ve belirlenmiş bir yerde durulması-gelişmekte olan hücrelerin özellikleridir. En azından gözlenen üç mekanizmanın, kullanılan ve etrafa yayılan kimyasal maddeleri, hücre yapışma moleküllerini (CAM’lar) ve dokunma çıkıntılarını içerdiği görülür.
 Yayılan kimyasal maddelerin gradientleri, sinir hücrelerinin kimyasalların kaynağına doğru ameboyit bir tarzda hareket etmelerine yardımcı olup, nöral krest hücrelerinin “aşağıya”, kortikal hücrelerin dışarıya doğru ve diğer hücrelerin ön (anteriyör), arka (posteriyör) ya da sırt (dorsal), karın (ventral) yönünde hareketlerine kılavuzluk ederler. Ancak hücrelerin izlediği yolların çoğu CAM’lar ve duyu çıkıntılarının bulunduğu diğer iki mekanizmayı içermektedir. Nöral krest hücreleri (omuriliğin etrafına ulaşan ve notokordu geçen glikoprotein filamentlerince yol gösterilen) ve kortikal hücreler (korteks boyunca fılamentlerin merkezden çevreye doğru ışınsal hareketlerini izleyen) CAM’ların özel oranları yardımıyla ayrı ayrı yollarını belirgin bir şekilde yeniden düzenlerler. Kendilerine rehberlik eden hücreleri kısmen çevrelerler daha sonra onların yanında yakın dokunma ilişkisini sürdürerek, hem kendisinden hem de substrattan yapılan CAM’lar arasında daha iyi eşleşmeler bularak hareket ederler. Göç eden bir hücre değdiği hücre üzerinde en uygun CAM haberleşmesiyle karşı karşıya kaldığı zaman hareketini durdurur ve yerleşeceği yere kendini tutturacak olan hücre-hücre bağlantılarını oluşturmaya başlar. Sinir hücresi yüzey belirleyicilerinin moleküler özgüllüğü öyle kusursuzdur ki her nöron sınıfının (iki hücre kadar az sayıda hücre bir sınıf yapar) zarında farklı molekülleri vardır; bu CAM’lardan daha özgül olan yüzey kimyasallarının nöronların kendilerine özgü bir ağ oluşturmasında son evresinde yer aldığı anlamına gelir.
Yayılan kimyasal maddelerin gradientleri, sinir hücrelerinin kimyasalların kaynağına doğru ameboyit bir tarzda hareket etmelerine yardımcı olup, nöral krest hücrelerinin “aşağıya”, kortikal hücrelerin dışarıya doğru ve diğer hücrelerin ön (anteriyör), arka (posteriyör) ya da sırt (dorsal), karın (ventral) yönünde hareketlerine kılavuzluk ederler. Ancak hücrelerin izlediği yolların çoğu CAM’lar ve duyu çıkıntılarının bulunduğu diğer iki mekanizmayı içermektedir. Nöral krest hücreleri (omuriliğin etrafına ulaşan ve notokordu geçen glikoprotein filamentlerince yol gösterilen) ve kortikal hücreler (korteks boyunca fılamentlerin merkezden çevreye doğru ışınsal hareketlerini izleyen) CAM’ların özel oranları yardımıyla ayrı ayrı yollarını belirgin bir şekilde yeniden düzenlerler. Kendilerine rehberlik eden hücreleri kısmen çevrelerler daha sonra onların yanında yakın dokunma ilişkisini sürdürerek, hem kendisinden hem de substrattan yapılan CAM’lar arasında daha iyi eşleşmeler bularak hareket ederler. Göç eden bir hücre değdiği hücre üzerinde en uygun CAM haberleşmesiyle karşı karşıya kaldığı zaman hareketini durdurur ve yerleşeceği yere kendini tutturacak olan hücre-hücre bağlantılarını oluşturmaya başlar. Sinir hücresi yüzey belirleyicilerinin moleküler özgüllüğü öyle kusursuzdur ki her nöron sınıfının (iki hücre kadar az sayıda hücre bir sınıf yapar) zarında farklı molekülleri vardır; bu CAM’lardan daha özgül olan yüzey kimyasallarının nöronların kendilerine özgü bir ağ oluşturmasında son evresinde yer aldığı anlamına gelir.
![Sinir Hücrelerinin Göçü]() Akson ve Sinapsların Oluşumu
Akson ve Sinapsların Oluşumu

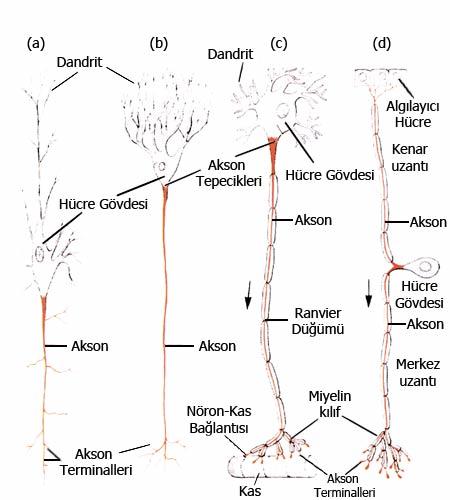
Bir sinir hücresi sinir sistemindeki daimi yerine ulaştığında, aksonlarını (bilgi iletimi için özelleşmiş uzun ince uzantılar) özel hedef hücrelere göndermelidir. Burada yeniden hem kimyasal hem de dokunumsal bilgilerin bir rol oynadığı görülür. Gelişen aksonun uzayan ucu “büyüme konisi” olarak bilinen nadir bir şekil gösterir.
Bu yapı ilk olarak yaklaşık bir asır önce İspanyol histolog Santiago Ramony Cajal tarafından tanımlanmıştır. Büyüme konisi sürekli olarak uzar ve bazı belirgin rehber hücrelerin varlığı ve özel kimyasallar için bir çevre oluşturmak için filopodia denen dikenimsi yalancıayaklarını çıkartır. Koni, bir rehber hücrenin (genellikle diğer bir sinirin aksonu) kimyasal ya da dokunumsal uyarılarıyla karşılaşırsa kısmen onu sarar ve onun boyunca uzar.
İlk olarak Kaliforniya Teknoloji Enstitüsünden Roger Sperry tarafından ileri sürülen, kimyasalın “ilk adım” stratejisi için kanıt, özellikle aksonların büyümesindeki zorlamadır. Pek çok sinir hücresi, hedeflerine ulaşır ulaşmaz aksonlarını, dolaylı olarak; fakat önceden belirlenen yönlerde hedef hücrelere gönderirler. Arasından aksonların geçtiği hücresel “özel alanın” yeniden düzenlenmesi hepsi için eşit olacak şekilde yeniden rotalanmaya neden olabilir. Örnegin, farelerde beyinin bir kısmını n görmeyle ilgili alanından çıkan aksonlar görmeyle ilgili korteksin 4. tabakasına uzanırlar. Aksonların, 4. tabakadaki özel olarak belirlenen hücreleriyle karşılaşıncaya kadar korteksin daha alt tabakaları arasından radyal (ışınsal bir tarzda) olarak basit bir şekilde büyüdüğünü tahmin edebiliriz; fakat bu böyle olmaz. Hayvanların sersemleyerek yürümesine neden olan mutasyonlu fare suşlarında kortikal tabakalar ters konumda olup, aksonlar ters konumlu korteks içinde büyüyüp, 6. tabakadaki bazı esas kimyasal belirleyicilerle karşılaşıncaya kadar 4. tabakadaki hedeflerine doğru sürekli ilerlerler ve daha sonra geriye dönüp geriye doğru göç edip 4. tabakayı bulurlar. Bu durum, bu aksonların korteksin önce 6. tabakasını bulmaya daha sonra da 4. tabakaya hareket etmesi gerektiğine göre programlandırıldığını gösterir.
 Omurgalılarda, bir milyon kadar çok sayıda akson korteksin görme, duyma ve dokunumsal alanlarında bulunan hedef hücrelerin büyük uzantılarından birine vücuttan bilgi gönderebilir. Bu akson hedef hücre bağlantıları uzamsal düzenlenmiş olup genetik olarak önceden belirlenmiştir. Görmeyle ilgili sistemde bulunan 10^6 özel moleküler işaretin tüm aksonların doğru hedef hücreleri bulmasını sağlaması olası görünmemektedir. Büyük uzantılarından birine vücuttan bilgi gönderebilirler. Basit bir ikikoordinatlı morfogen gradientinin kafi derecede yeterli olduğunu düşünmek de akla yatkın değildir.
Omurgalılarda, bir milyon kadar çok sayıda akson korteksin görme, duyma ve dokunumsal alanlarında bulunan hedef hücrelerin büyük uzantılarından birine vücuttan bilgi gönderebilir. Bu akson hedef hücre bağlantıları uzamsal düzenlenmiş olup genetik olarak önceden belirlenmiştir. Görmeyle ilgili sistemde bulunan 10^6 özel moleküler işaretin tüm aksonların doğru hedef hücreleri bulmasını sağlaması olası görünmemektedir. Büyük uzantılarından birine vücuttan bilgi gönderebilirler. Basit bir ikikoordinatlı morfogen gradientinin kafi derecede yeterli olduğunu düşünmek de akla yatkın değildir.
Pek çok araştırmacı, başlangıçtaki dallanmaların tam olmadığına ve son şeklin dinamik bir şekilde sinirsel rekabet denilen bir olay aracılığıyla belirlendiğine inanırlar.
Kaynakça:
https://www.mananatomy.com
Yazar: Taner Tunç