
 Yaşam, kararlılık ve değişim arasındaki optimal bir denge üzerinden işlemektedir. Genetik mesaj güvenilir biçimde devam ettirilirken, aynı zamanda, özellikle rekabet, avlanma, habitat ve iklim bakımlarından önceden tahmin edilemeyen değişimler karşısında doğal seçilimin uygun genleri ortaya çıkarması için gerekli olan varyasyon da sağlanır. Eşeysel organizmalar eşleşmede (eşeysel rekombinasyon) kullanacakları haployit gametler oluşturmak için çok güç harcarlar. Diployit yavru dölün eşeysiz klonlanması metabolik olarak daha az israflıdır ve gamet üreten hücreler krossing-over gerçekleştirebilmek için büyük çaba harcarlar. Pek çok araştırmacı, bu iki mekanizmanın kararlılık ya da değişim işlevini gerçekleştirdiğine inanmakta ancak hangisinin temel olduğu konusunda anlaşamamaktadır. Rekombinasyon ve krossing overin mekanizmaları ve sonuçları, varyasyonu üretme ve sınırlama gücünü tanımlamaktır.
Yaşam, kararlılık ve değişim arasındaki optimal bir denge üzerinden işlemektedir. Genetik mesaj güvenilir biçimde devam ettirilirken, aynı zamanda, özellikle rekabet, avlanma, habitat ve iklim bakımlarından önceden tahmin edilemeyen değişimler karşısında doğal seçilimin uygun genleri ortaya çıkarması için gerekli olan varyasyon da sağlanır. Eşeysel organizmalar eşleşmede (eşeysel rekombinasyon) kullanacakları haployit gametler oluşturmak için çok güç harcarlar. Diployit yavru dölün eşeysiz klonlanması metabolik olarak daha az israflıdır ve gamet üreten hücreler krossing-over gerçekleştirebilmek için büyük çaba harcarlar. Pek çok araştırmacı, bu iki mekanizmanın kararlılık ya da değişim işlevini gerçekleştirdiğine inanmakta ancak hangisinin temel olduğu konusunda anlaşamamaktadır. Rekombinasyon ve krossing overin mekanizmaları ve sonuçları, varyasyonu üretme ve sınırlama gücünü tanımlamaktır.
Varyasyon
 Eşeysel üremenin en önemli sonucu, yeni karakter kombinasyonlarına sahip yavrular meydana getirilmesidir. Homolog kromozom çiftindeki her üye farklı bir ebeveynden gelir ve aynı tür RNA, yapısal proteinleri ve enzimleri kodlayan genler içermelerine karşın iki homolog gendeki baz dizileri birbirinin aynı değildir. Aslında, bir organizmanın babasından kalıtılan bir gen kopyası anneden kalıtılan gen kopyasından en azından küçük oranda farklıdır.
Eşeysel üremenin en önemli sonucu, yeni karakter kombinasyonlarına sahip yavrular meydana getirilmesidir. Homolog kromozom çiftindeki her üye farklı bir ebeveynden gelir ve aynı tür RNA, yapısal proteinleri ve enzimleri kodlayan genler içermelerine karşın iki homolog gendeki baz dizileri birbirinin aynı değildir. Aslında, bir organizmanın babasından kalıtılan bir gen kopyası anneden kalıtılan gen kopyasından en azından küçük oranda farklıdır.
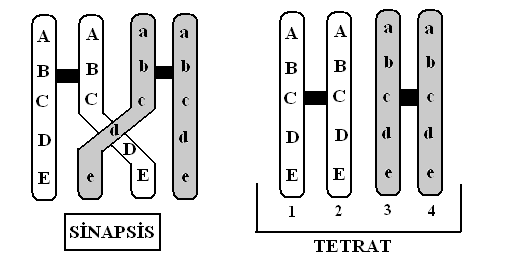
Aynı genin farklı biçimlerine aleller denir ve bunların ürettiği yapısal proteinler ya da enzimler birbirinden biraz farklı etkinliğe sahiptirler. Örneğin, göz pigment genlerinin her iki kopyasının da bozuk, renksiz tarayıcı pigmentleri kodladığı insanlarda mavi renkli gözler gelişir; kahverengi gözlüler işlevsel bir pigment kodlayan genlerden en az bir kopyaya sahiptirler. Bir organizma iki farklı ebeveynden gelen alellerin bir karışımı olduğu için kromozomlarının her ebeveyeninkinden farklı alleller içermesi hemen hemen kesindir ve onun morfolojik, fizyolojik ve davranışsal pek çok özelliği, buna uygun biçimde, farklı olacaktır. Varyasyonun neden eşeysel rekombinasyonda gerçekleştiğini anlamak için altı kromozomlu bir hipotetik organizmayı düşünelim. Uç kromozom erkek ebeveynin gametinden (mayoz sonucu), üç kromozom ise dişi ebeveynin gametinden gelerek bu diployit organizmada üç homolog çift meydana getirmişlerdir. Üreme için gamet meydana getiren bir bireyde gerçekleşen her mayozda, bu organizmanın tüm anna kökenli kromozomları bir gamete, tüm baba kökenli kromozomları başka bir gamete gidebilir, ancak diğer tüm kombinasyonlar da eşit biçimde olasıdır. Bir organizma üç kromozom çiftine sahipse, gametlerde sekiz farklı kombinasyon bulunabilir. İki gametin zigot oluşturduğu böyle bir türde 64(8×8) farklı kromozom kombinasyonu söz konusu olacaktır. İnsanlar açısından, olağandışı çokluktaki kromozomlarımız düşünüldüğünde, aynı anne babadan gelen yavrulardaki olası kromozom kombinasyonlarını n sayısı 7×1013’tür.
Krosing-over olmasa bile, basit eşeysel rekombinasyon önemli oranda varyasyon meydana getirir ve bu durum, mayozun karmaşık iki evreli süreci yerine daha basit bir süreç geçseydi dahi söz konusu olurdu: mitozun telofaz evresini sona erdirdiğini gösteren tek kromatitli homolog kromozomlar, teorik olarak, iki haployit hücreye dağılabilirlerdi. Mayoz 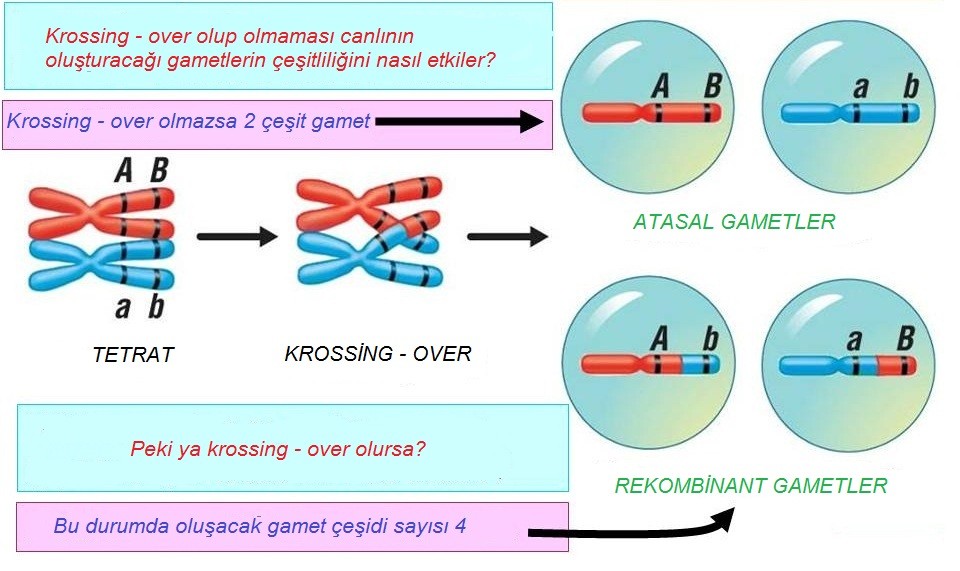 öncesindeki DNA replikasyonu ve tüm ikinci bölünme aşaması, bu nedenle gereksizleşirdi. Ancak bu kestirme yol, ekonomik olmasına karşın, organizmayı, bir ikinci yeni kombinasyonlar üretme yolu olan krossing-overin sağladığı yararlardan yoksun bırakırdı.
öncesindeki DNA replikasyonu ve tüm ikinci bölünme aşaması, bu nedenle gereksizleşirdi. Ancak bu kestirme yol, ekonomik olmasına karşın, organizmayı, bir ikinci yeni kombinasyonlar üretme yolu olan krossing-overin sağladığı yararlardan yoksun bırakırdı.
Oldukça düzenli bir süreç olan krossing-over değişken yapıya sahip olası gametlerin sayısını astronomik biçimde artırır. Bunun nedeni, krossing-overin tamamen rastgele noktalardan gerçekleşmesi ve istisnasız her hibrit kromozomun öbürlerine benzemeyen noktalardan olan kopmalarla meydana getirilmesidir. Eğer yalnızca iki kromozomlu ve genleri binlerle ifade edilen bir örneğe odaklanacak olursak, mayoz sonrasında genetik bilginin başında geçenlere dair bir fikir edinebiliriz. Bu durumda, varsayalım ki, bir kromozomun bir ucundaki tek bir gen hayvanın post rengini, aynı kromozomun diğer ucundaki bir gen ise vücut büyüklüğünü kontrol etsin.
Babanın siyah post (F diyebiliriz) kodlayan bir geni ve büyük vücut olmasını kodlayan bir geni ( S) verdiğini, anneden gelen alellerin gri post (f) ve küçük vücut (s) biçiminde, farklı olduğunu varsayalım. Babadan gelen F ve S’nin; annede ki f ve s gibi, hücre mitoz geçirirken birlikte (ya da bağlı) kalmalarına karşın, mayoz sırasındaki krosing over yeniden düzenlenmeler üretecektir. Gerçekte, krossing over genellikle bir kaç bölgede gerçekleştiği ve herhangi iki homolog kromozomla ilgili olduğu için, bir atasal kromozomun dokunulmamış biçimde mayozdan çıkması olasılığı, yüzde 50’nin oldukça altındadır. Dahası, krossing over bir gen içinde dahi gerçekleşebilir; yeni aleller dahi genin kopyalarından farklı olmaları durumunda, yapılabilir.
![Krossing-Over Olayının Canlılar İçin Önemi]() Kararlılık
Kararlılık
 Kararlılık
KararlılıkBuraya kadar rekombinasyon ve krossing-overin nasıl genetik yenilik ürettiğinden oldukça söz ettik. Peki, diğer alternatif, kararlılığın devamı, ne anlama gelir? Değişimsel bir argüman, onarılmayan mutasyonların bir genomda, bazı genlerin her iki kopyasının da hasarlı ya da işlevsiz oluncaya dek yavaş yavaş birikeceğini iddia eder. Sonuç olarak, kuşaklar geçtikçe, bir organizmadan türeyen yavrular zayıflar, buna karşın, daha çok mutasyona maruz kalırlar. Böyle genlerin işlevsel kopyalara sahip olmalarının iyi bir yolu rekombinasyon geçirmektir. Döllenmiş bir yumurta olasılıkla her genin en azından bir tane iyi kopyasına sahiptir. Çünkü akraba olmayan bireylerden gelen iki gametin aynı mutasyonlara sahip olması olanaklı değildir. Eşey, o halde kararlılığın devamını sağlamak için varolabilir.
Krossing overi koruyucu gören argüman aşağı yukarı aynı yolu izler. Bazen gerçekleştiği gibi bir kromozomun DNA’sının her iki iplikçiği de hasar gördüğünde onarım enzimleri sorunu çözemez çünkü kalıp olarak kullanılabilecek iyi bir iplikçik yoktur. Ancak homolog kromozomda genin tam bir kopyası vardır. Bazı araştırıcılar krossing overin çift iplikcikli hasar bölgeleri yakınında gerçekleştiğine ve böylece homolog kromozomdaki aynı bölgeden kopya çıkarmakla onarım enzimlerinin hasarlı kromozomu düzelttiğine inanmaktadırlar.
Son olarak, rekombinasyon ve krossing-overin zıt amaçlara hizmet için değişmiş olmaları ya da bu iki amacı aynı anda gerçekleştiriyor olmaları bütünüyle olasıdır. Bunun nedeni ciddi mutasyonların onarımı, telafisi aynı zamanda büyük miktarlarda çeşitlilik meydana getirilmesidir. Moleküler ve değişimci biyologları biraraya getiren pek çok konudan biri olan bu önemli problem çözülmeyi beklemektedir.
Kaynakça:
Khan Academy
Yazar: Taner Tunç