
 Yüksek bitkiler, özelleşmiş dokulara (kökler, gövdeler, yapraklar ve üreme organları) sahiptirler. Eğer bir bitki çimlenir, büyür ve ürerse, bu dokuların aktivitelerinin koordine edilmesi gerekir.
Yüksek bitkiler, özelleşmiş dokulara (kökler, gövdeler, yapraklar ve üreme organları) sahiptirler. Eğer bir bitki çimlenir, büyür ve ürerse, bu dokuların aktivitelerinin koordine edilmesi gerekir.
Gövde ve köklerde büyümenin yönlendirilmesi ve düzenlenmesinde, yaprakların üretimi ve dökülmesinin zamanlanmasında, tohum çimlenmesinin başlatılmasında ve diğer pek çok işlevin düzenlenmesinde kimyasal kontrol, rol oynayabilir. Bir kimyasal, üretildiği özelleşmiş dokulardan salgılandıktan sonra bitkinin diğer kısımlarına taşınır (genellikle organizmanın içsel taşınım sistemiyle) ve bir dizi özgün, hedef hücrede bir etki yaratırsa, bu madde hormon tanımına uyar.
Bitkisel hormonlar, en azından bilinenler, gövde ve kökün apikal meristemlerinde, büyümesini sürdüren genç yapraklarda ya da tohumlar ya da meyveler gibi bitkinin aktif olarak büyüyen kısımlarında en fazla üretilirler. Bu hormonları oluşturan dokular, sıklıkla da meristem dokularının kendileri, hormon üretimi için özelleşmiş olmalarına karşın, hayvanlardaki hormon üreten dokular kadar özelleşmemişlerdir. Bitkilerde, yüksek hayvanlardaki endokrin bezlerine eşdeğer hormon üreten organlar bulunmaz.
Oksinler
Tümü oksin olarak isimlendirilen ve çok çalışılmış bir grup bitki hormonu, farklı bitki dokularının büyümesi üzerinde şaşırtıcı çeşitlilikte etkiler gösterir. Esas olarak, bitkiler, hem apikal meristemin bulunduğu bölgelerde gerçekleşen hücre bölünmesiyle hem de özellikle gövde dokusunda daha önceden mevcut hücrelerin uzaması ve genişlemesiyle büyürler. Daha önce değindiğimiz gibi oksinler hücre uzamasını başlatırlar.
Oksinler ve Gövdelerde Fototropizma
Pek çok bitkinin şiddetli bir biçimde ışığa doğru yönelme eğiliminde olduğunu biliriz. Oturma odasında bir saksıda yetişen bir bitki pencere yönünde büyür; bu bitkiyi döndürdüğünüzde odadaki insanlara daha iyi görünecektir; ancak kısa bir süre sonra gövdenin yeniden penceredeki ışığa doğru yöneldiğini görürsünüz. Işığa sonucu tepki olarak farklı büyüme sonucu ortaya çıkan bu olay, Yunanca “ışık” ve “dönüş” kelimelerinden kökenlenen fototropizma olarak adlandırılır. Diğer tropizmalar, büyümede, diğer uyarılara karşı oluşan tepkiyi kapsar. Örneğin, gravitropizma, yerçekimine karşı gösterilen bir tepkidir. 
Bitki gövdelerinde fototropizma uyarının yönüne doğru, yani pozitiftir. Kökler ise ışık uyarısından uzaklaşan bir büyüme, yani negatif fototropizma gösterirler. Bitkilerde fototropizmaya ilişkin ilk çalışmalar Charles Darwin ve oğlu Francis tarafından yaklaşık 1880’de yapılmıştır. Bu araştırıcılar daha sonrakiler gibi denemelerini çimler ve bunlar akraba genç bitkilerin ilk yapraklarını saran silindirik bir kın üzerinde gerçekleştirmişlerdir. Koleoptil olarak adlandırılan bu kın, esas olarak hücre uzaması ile büyür ve ışığa doğru yönelerek çok kuvvetli bir pozitif fototropik tepki gösterir. Kıvrılma, büyümede oluşan farklılık ile başarılır: Koleoptilin ışık olmayan tarafındaki hücreler uzayarak, uç’u ışığa doğru yönelmeye zorlarlar.
Darwinler, koleoptilin ucunun küçük bir siyah şapka ile kapatılması halinde bitkinin ışığa yönelemediğini göstermişlerdir. Buna karşılık, uçları kapatılmamış ya da ışık geçiren bir örtü geçirilmiş kontrol koleoptiller ise, beklendiği gibi, ışığa yönelmişlerdir. Uç kısmın değil de koleoptilin kaidesinin ışık alması engellendiğinde kıvrılmanın önlenemediği gözlenmiştir. Böylece, koleoptilin ucunun ışığın algılanması da kilit rol oynadığı ortaya çıkmıştır. Daha sonra, bu bilginin, fototropik tepkinin -farklı büyümeye bağlı olarak kıvrım oluşması- başarıldığı büyüme bölgesine (uç kısmın hemen altındaki) geçirilmesi gerekir. Bu, ucu kesilen koleoptillerin kıvrım yapamadıklarının bulunmasıyla Darwinler tarafından kanıtlanmıştır.
Uçları kesilmeyen kontrol bitkilere ait koleoptillerin başka zararlar görmeleri halinde bile, normal olarak kıvrıldıkları görülmüştür. Fototropik tepkinin durmasında yaralanmaya karşı oluşan bir reaksiyonun değil, ucun yokluğunun neden olduğu açık olarak anlaşılmıştır. Darwinler, ışığın, koleoptilin ucu tarafından algılandığı ve yaratılan etkinin daha sonra üst taraftan kıvrımın oluşacağı alt tarafa geçirildiği sonucuna varmışlardır.
Yaklaşık 30 yıl sonra Danimarka’da P. Boysen-Jensen, Darwinlerin öne sürdükleri “etkinin” elektriksel ya da sinirsel değil, olasılıkla kimyasal olduğu yolunda ilk kanıtları elde etmiştir. Bu araştırıcı, yulaf koleoptillerinin ucunu kesmiş, her bir kesik ucun üstüne ince bir jelatin tabaka yerleştirmiş ve daha sonra kesik ucu jelatin tabakanın üzerinde konumlandırmıştır. Böylece, kesik uç koleoptilin geri kalan kısmından ince bir jelatin tabakayla ayrılmıştır. Daha sonra, uç kısım yandan ışıklandı rıldığında, koleoptilin kaidesi ışığa yönelmiştir. Uç, ışık uyarısını almış ve uçtan bir mesaj jelatin engeli geçerek kaidede kıvrılma oluşturmuştur. Bu deney, bir elektriksel ya da sinirsel mesaj olasılığını tamamen devre dışı bırakmamışsa da, böyle bir olasılığı önemli ölçüde zayıflatmış ve difüzyon yapabilen bir kimyasalın iş gördüğünü kanıtlamıştır.
O uc’un, karanlıkta bile, bir yulaf koleoptil kaidesinin kıvrılmasına neden olabildiği Macaristan’da, 1918’de A. Paal tarafından gösterilmiştir. Bu araştırıcı, ucu kestikten sonra, sütunun üzerinde merkezden uzak bir yere yerleştirmiştir. Karanlıkta, ucu, sütunun sağ tarafına koyduğunda, koleoptil sola kıvrılmıştır. Ucu sol tarafa yerleştirdiğinde ise, koleoptil sağa kıvrılmıştır. Uç kısmın altında bulunan koleoptil parçası, ucun altında bulunmayan kısımdan çok daha hızlı büyümüştür. Koleoptilin gösterdiği bu asimetrik uzama, onun, daha fazla uzayan tarafın aksi yönünde kıvrılmasına neden olmuştur. Ucun, koleoptilin gövdesinden sürekli olarak aşağı doğru hareket eden ve daha alttaki hücrelerin doğrudan büyümesine neden olan bir madde üretmiş olması, mantıklı kabul edilebilirdi.
Bu, büyüme bölgesi ucun altında olsa bile, uçsuz bir koleoptilin büyüyemiyeceğine ilişkin şaşırtıcı gözlemi açıklayabilecekti. Bu yorumun ışığında değerlendirilen Boysen Jensen’in çalışması, yandan gelen ışığın, gövdenin her iki yanından aşağıya taşınan bu hipotetik maddenin nispi miktarını değiştirdiği ve dolayısıyla bir tarafın diğerinden daha fazla büyümesine neden olduğunu düşündürüyordu.
Uçtan aşağı doğru taşınan büyüme uyarısını bir kimyasal olduğunu gösteren deneyler, daha sonra 1926’da, Hollanda’da Fritis Went tarafından rapor edildi. Araştırıcı, yulaf koleoptillerinin ucunu kesti ve kaideleri aşağıya gelecek şekilde bu izole uçları yaklaşık bir saat süre ile agar blokların üzerine yerleştirdi (Deniz yosunlarından elde edilen ve jelatinimsi bir madde olan agar, laboratuvarda genellikle kültür ortamı olarak kullanılır). Daha sonra agar blokları uçsuz koleoptil sütunların üzerine koydu. Uçları kesik koleoptil parçaları bir süre büyümeye bırakıldılar. Agar blokları merkezden uzakta konumlandırıldıklarında, koleoptiller karanlıkta bile kıvrılabiliyorlardı. Kontrol olarak kullanılan agar blokları, bu etkilerin hiç birini oluşturmadı. Uç kısımlarda büyümeyi teşvik edici bir maddenin yine bu kısımlarda sentezlendiği ve oradan üzerlerinde uçların oturduğu agar bloklara geçtiği açıktı. Daha sonra, bu bloklar koleoptil sütunların üzerine yerleştirildiklerinde, kimyasal, onlardan koleoptil sütunlarına taşınıyor ve uzamayı teşvik ediyordu. Bu deney, uyarının elektriksel ya da sinirsel olma olasılığını ortadan kaldırdı. Went, difüzyonla geçebilen bu kuramsal hormona auxin (Yunanca “büyümek” kelimesinden türevlenmiştir.) adını verdi. Bugün de biyologlar öksinleri Went’in deneyini esas olarak tanımlamaktadırlar: eğer araştırılan maddeyi içeren bir agar blok, karanlıkta kesik ucun bir tarafına yerleştirildiğinde, ucu kesik yulaf koleoptillerinin kıvrılmasına neden oluyorsa, bu madde bir öksin olarak sınıflandırılmaktadır.
Bazıları bitkilerde doğal olarak bulunan, diğerleri ise yalnızca laboratuvarlarda sentezlenen pek çok kimyasal, Went’in testinden geçmiş ve bunlara genel olarak öksin adı verilmiştir. Çok sayıda doğal kaynaktan izole edilmiş olan indolasetik asit en fazla araştırılmıştır. Indolasetik asitin başlıca aktif, temel doğal öksin olduğuna ilişkin kanıtlar vardır. Diğer doğal bileşiklerde gözlenen öksin etkenliği bunların bitki tarafından indolasetik asite dönüştürülmesinin sonucu olabilir.
Tartıştığımız deneyler, koleoptilin ucunun, yukarıdan aşağıya doğru taşınan ve koleoptilde hücre uzamasını teşvik eden öksininin koleoptilin ucu tarafından üretildiğini göstermiştir. Paal ve Went tarafından elde edilen sonuçların da gösterdiği gibi, uçtan salındıktan sonra, öksinin normal olarak yanal hareketi azdır; hormon, yalnızca doğrudan serbest bırakılma noktasının altındaki hücrelere ulaşır ve teşvik eder. Kıvrılma oluşturan büyüme farklılığı bitki tek yandan ışıklandırıldığında ortaya çıkar. Bitkinin ışık alan tarafında oksin miktarı azalan. Bunun sonucunda, ışık alan taraf az ışık alan taraftan daha yavaş büyür. Bu asimetrik büyüme sonucu yavaş büyüyen, ışık alan tarafa doğru kıvrılma oluşur.
Bitki fizyologları, uç tarafın ışığı nasıl algıladığını henüz kesin olarak bilmemektedirler. Bu tepki yalnızca mavi ışı k tarafından başlatılmaktadır. (Hatırlanacağı üzere, beyaz ışık tüm renklerin bir karışımıdır.) Bu, algılayıcı pigmentin, turuncu (maviyi absorblayan herhangi bir maddenin gözle görülebilir rengi) olması anlamına gelir. Fakat algılayıcı pigment olma olasılığı taşıyan tüm renklerin bu özellikte olmadıkları kesinlikle ispatlanmış olup, ilgili pigment henüz bilinmemektedir.
Ayrıca bitki fizyologları ışık algılanmasının asimetrik oksin dağılımı ile nasıl eşleştiğini de bilmemektedirler. Kanıtlar, uçta, ışık alan taraftan ışık almayan tarafa doğru aktif yanal taşınımının olduğunu göstermekle birlikte; bu konu halen tartışılmaktadır.
Oksin Etkisinin Hücresel Temeli
Gövde kıvrılmasının oksine bağlı olduğunun anlaşılmasından sonra, çalışmalar, bilgi akışındaki üç evre üzerinde odaklaşmıştır: ışık nasıl algılanmakta, oksinin taşınımı nasıl kontrol edilmekte ve oksin gerçekte kıvrılmaya nasıl neden olmaktadır.
Oksinin çeşitli işlevlerinden her birinin nasıl gerçekleştirildiği konusunda öğrenilecek hâlâ pek çok şey bulunmasına karşın, bu yöndeki gelişmeler hızlı olmuştur. Büyümekte olan uçlarda, oksin, sürekli olarak üretildiğinden, ışık reseptörlerinin bu hormonun asimetrik dağılımını başlatarak ışığa tepki gösterdikleri açıktır. Reseptör moleküller, ışığa tepki olarak, hormonu, açık bir biçimde gövdenin ışık almayan tarafına doğru yönlendirmektedir.
Yatay ve dikey oksin taşınımıda olasılıkla, benzer süreçleri kapsamaktadır. İletim sisteminden bağımsız hücreden hücreye taşınımın çok hızlı olması, aktif taşınım işaret etmektedir. Kontsantrasyon gradientine karşı taşınabilmesine karşın, metabolik engelleyiciler bulunduğunda, oksinin taşınamadığının gösterilmesi bu görüşü desteklemektedir.
Asit-büyüme hipotezi olarak bilinen bir modele göre, oksin, gövdede büyümekte olan hücrelere ulaşınca, sitoplazmadan mebranın dışındaki hücre çeperine H+ iyonlarını taşıyan pompaları etkinleştirmektedir.
Buradaki pH, hücre çeperindeki selüloz fibrillerin arasındaki hidrojen bağlarının parçalanmasına neden olmaktadır. Sonuç olarak, çeperin esnekliği armakta ve hücreler daha fazla su alarak uzayabilmektedirler. Gövdenin ışık almayan tarafında daha fazla oksin bulunduğundan, oradaki hücreler ışık alan taraftaki hücrelerden daha fazla uzarlar ve gövde kıvrılır. Büyüme bölgesi dışındaki hücreler oksine duyarsızdırlar.
Oksinler ve Gövdelerde Gravitropizma
Bitkiler yanlızca ışığa değil, yerçekimine de tepki gösterirler. Eğer saksıdaki bir bitkiyi karanlıkta yan yatırıp birkaç saat o konumda bırakırsanız gövdenin yukarı doğru kıvrıldığını görürsünüz. Bu bir negatif gravitropik (ya da “geotropik”) tepkidir: gövde, yerçekiminden uzaklaşarak yukarı doğru kıvrılmaktadır. Oksinin bu tür bir bitki büyüme tepkisini kontrol edebileceğini varsaymak mantıklı görünmektedir.
Hollanda’da Herman Dolk, yatay olarak konumlandırılmış bir gövdede, oksin konsantrasyonunun, alt tarafta artarken üst tarafta azaldığını göstermiştir. Bu eşit olmayan oksin dağılımın alt taraftaki hücrelerin üst taraftaki hücrelerden daha hızlı uzamalarını teşvik etmesi ve bu nedenle gövdenin yukarı doğru dönmesi gerekir. Bununla birlikte, konu o denli basit değildir: oksin dikotillerin gravitropizmasına katılmayabilir ve monokotillerde ise en azından diğer bazı hormonlardan yardım alabilir.
Köklerdeki durum da belirgin değildir. Tartışmalı olmasına karşın, kök şapkasındaki amiloplastlar olarak adlandırılan özelleşmiş nişasta plastitlerinin dağılımlarının temel bir rol oynama olasılığı vardır. Amiloplastlar (gövdede ve meristem hücrelerinde de bulunurlar) Ca++ iyonlarının serbest kalmasını tetikleyerek süratle kök şapkası hücrelerinin kaidesine yerleşirler. Kalsiyumun ise kök şapkasının arkasındaki büyümekte olan hücrelerin üst ve alt zarları arasında büyüme hormonlarının asimetrik dağılımına neden olduğu sanılmaktadır. Oksinin, açıkça bu reaksiyonda taşınan maddelerden biri olmasına karşın, büyümenin yönlendirilmesin de temel bir rol oynadığına ilişkin yeterince kanı yoktur.
Oksinler ve Yanal Tomurcuklarm Engellenmesi
Tepe tomurcuğunda üretilen oksin, gövdeden aşağı doğru taşınarak ana gövdenin uzamasını teşvik ederken yanal tomurcukların gelişimi engeller. Bunun sonucunda, tepe tomurcuğu, gövdenin diğer kısımları üzerinde apikal dominansi oluşturur. Böylelikle bitkinin büyüme için bitkinin enerjisinin ana gövdeye gitmesi ve yan dalları nispeten kısa olan uzun boylu bir bitkinin oluşması sağlanır. Uzun dallar, apikal dominansinin etkisinin az olduğu, tepe tomurcuğundan yeterince uzaktaki tomurcuklardan gelişir. Bununla birlikte, terminal uç uzaklaştırılırsa, apikal dominansi geçici olarak ortadan kalkar ve üst taraftaki yanal tomurcukların bazıları dallar oluşturarak büyümeye başlar. Bu dalların tepe tomurcukları daha alttaki herhangi bir tomurcuk üzerinde kısa sürede baskınlık oluşturur. Çiçek ve ağaç yetiştiricileri, pek çok çiçeklenme noktası bulunan çalımsı görünümlü, iyi dallanmış bitkiler üretmek için, her mevsim bir ya da daha fazla olmak üzere, sıklıkla, yetiştirdikleri bitkilerin tepe tomurcuklarını budarlar.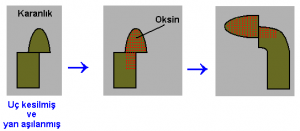
Bununla birlikte, yanal tomurcukların tepe tomurcuğu tarafından değilde, genç yapraklar tarafından kontrol altında tutulduğu bitkilerde tepe tomurcuğunun uzaklaştırılması iş görmez. Çeşitli tipteki bitkilerin farklı büyüme formları , türler arasında farklı apikal dominansi derecelerini yansıtır.
İki ya da daha fazla dal gelişmeye başlamışsa, bunlardan biri diğerini engellemez. Bir dalın tepe tomurcuğu tarafından salgılanan oksin çoğunlukla gövdede yukarıdan aşağıya doğru taşındığından, diğer dalın tepe tomurcuğuna önemli miktarda ulaşmaz. Tek yönlü bu taşınım, diğer pek çok (belki de tümü) hücre gibi taşınıma katılan hücrelerin de fizyolojik olarak polarize olduklarını –yani bunların uç kısımlarının birbirlerine göre önemli farklılıklar gösterir- ve bu nedenle, belirli kimyasalların taşınımının yalnızca bir yönde ortaya çıktığını göstermektedir. Bu durumda, en olası farklılık, üst ve alt membranlarındaki taşıyıcı protenlerin yöneliminde olmaktadır.
Oksinler ve Meyve Gelişimi
Normal gelişimi oksinlerin uyarıcı etkisine bağlı olan organ meyvedir. Meyve ovaryumdan ya da çiçek tablasında gelişir. Döllenme oluşmazsa, genellikle meyva gelişmez ve meyve yerine, meyve sapının kaidesinde absisyon tabakası olarak adlandırılan, ince çeperli hücrelerin oluşturduğu zayıf bir tabaka meydana gelir. Bu tabaka herhangi bir hafif dokunma ile parçalanır ve solmuş olan çiçek, ovaryumu ile birlikte yere düşer. Diğer taraftan, eğer döllenme gerçekleşirse, absisyon tabakası oluşmaz ve ovaryum hızla büyümeye başlar. Pek çok türde ovaryumun meyveye dönüşümü sırasında (ya da bazı bitkilerde reseptakulumun) ortaya çıkan bu hızlı büyüme dönemi, döllenmeyi sağlayan polen danelerinin serbest bıraktıkları öksin tarafından başlatılır. Bu tür bitkilerde meyvenin büyüme ve gelişmesinin sürmesi tohumlar tarafından üretilen öksinlerin uyarısına bağlıdır.
Oksinlerin meyve gelişimini teşvik edici ve absisyon tabakasının oluşumunu engelleyici rolünün anlaşılmasıyla çekirdeksiz meyve üretme olanağı doğmuştur. Az sayıdaki bitki, bazen doğal olarak tohumsuz meyve oluşturur. Bu durumda ovaryumun kendi dokuları (ya da ilişkili yapılar) meyvenin oluşumu için döllenmeye ve bunun sonucunda da tohum oluşumuna gerek kalmaksızın bol miktarda oksin üretirler. Domatesler, salatalıklar (hıyarlar), kabaklar ve incirler gibi normal olarak tohum oluşturan bitkilerin döllenmeleri engellenir ve bu bitkilere tohumların ürettikleri miktarda yapay oksin uygulanırsa, tohumsuz meyve oluştururlar. Bu yöndeki denemeler sonucunda, 1934’te Japonya’da S. Yasuda tohumsuz salatalık, 1936’da Michigan Üniversitesi’nde F.G. Gustafson ise çekirdeksiz domates üretmişlerdir. O zamandan beri, diğer pek çok bitkide çekirdeksiz meyve üretilmiştir. Bununla birlikte, bazı bitkilerde, özellikle de tek tohum içeren bitkilerde yapay oksin uygulamasıyla çekirdeksiz meyve üretimi çabaları henüz bilinmeyen nedenlerle başarısızlıkla sonuçlanmıştır.
Bunun nedeni, en azından üzümlerde başka bir hormonun (şu an tartışılacak olan giberellin) meyve gelişiminden sorumlu olmasıdır. Çilekler ve böğürtlenler gibi diğer bitkilerde, tohumsuz meyveler üretilmekle birlikte, normal olarak tohum sert kınlar (ovaryumdan köklenmiş) tarafından kuşatıldığında meyveyi yiyen biri onun tohumsuz olup olmadığını söyleyemez. 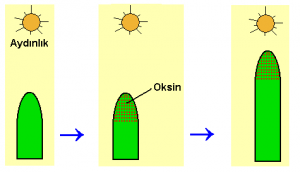
Meyve veren bitkilere oksin uygulamasının başka ticari uygulamaları da vardır. Meyve bağlanması için çoğunlukla normal tozlaşma gerekli olup bu meyvelerin irileşmesini sağlar. Bazı durumlarda oksin püskürtülerek meyvelerin boyutu arttırılabilir. Olgun meyvenin yere düşmesi de absisyon tabakasının oluşmasına bağlı olduğundan, olgunlaşan tohumlarda üretilen oksin giderek azaldığından olgunlaşma yaklaştığında, pek çok meyve ağacına oksin püsürtülmesi yaygın bir uygulama halini almıştır. Bu spreyler absisyon tabakası oluşumunu engelleyerek zamansız meyve dökülmesini azaltır (Zamansız dökülen meyvaların pek çoğu ticari değerini kaybeder).
Oksinler ve Yaprak Absisyonu
Döllenmemiş çiçekler bitkiden düşer. Çünkü çiçeklerde yüksek oksin üretimi olmadığında, çiçek sapının kaidesinde özel bir hücre tabakası, yani absisyon tabakası oluşur. Absisyon tabakasının oluşması sonucu, olgun meyve de yere düşer. Benzer şekilde, yaprak döken ağaç ve çalıların sonbaharda (ya da hastalanmış yaprakların yılın herhangi bir zamanında) yapraklarını dökme kısmen yaprak ayası tarafından üretilen oksin miktarındaki azalmanın bir sonucu olarak, genellikle (her zaman olmasa da) petiolün kaidesinde absisyon tabakası oluşmasına bağlıdır.
Kısacası, oksin, absisyonu engelleyici olarak iş görür; diğer bir hormon, yani etilen (kısaca tartışılacak), absisyonu artırıcı etki yapar.
Orta şiddetteki bir rüzgâr basıncı gibi herhangi bir hafif zorlama, absisyon tabakasında kopma başlatabilir. Çünkü absiyon bölgesindeki hücrelerin çeperleri, selülozu parçalayıcı bir enzim olan selülaz konsantrasyonundaki artış nedeniyle zayıflar.
Oksinler ve Hücre Bölünmesi
Oksinlerin hücre uzaması ve absisyon tabakası oluşumundaki rollerinin yanı sıra, belirli dokularda hücre bölünmesine katıldıkları da düşünülmektedir. Erken ilkbaharda, tomurcuklardan aşağıya doğru taşınarak kambiyumun etkinleşmesini sağlayan maddenin oksin olduğu açıktır. Böylece, yeni iletim dokuları oluşmaktadır. Sonbahar yaklaştıkça, tomurcuklar ve yapraklar tarafından üretilen oksin miktarındaki azalmaya bağlı olarak kambiyumun etkenliği de azalır.
Öksinler olasılıkla yan köklerin oluşumunu da başlatırlar. Daha önce gördüğümüz gibi, böyle kökler, genellikle endodermisin hemen iç kısmında, perisikl adı verilen nispeten farklılaşmamış hücre tabakalarından kökenlenirler. Perisikl hücreleri çoğu zaman meristematik etkenlik göstermezler. Bununla birlikte, perisikl’deki küçük bir hücre grubu, aralıklarla aktif bir biçimde bölünen meristematik dokuya dönüşerek, ana kökün dış kısmındaki dokulardan çıkarak toprağa giren yeni bir lateral kök oluşturur. Perisikl’da meristematik etkenlik başlatan uyarı öksinden gelir. Aslında, yan yana dallanma oluşturmak için bitki köklerine oksin uygulanabilir; ancak Oksinin hücre bölünmesini doğrudan teşvik edip etmediği belirsizdir; örneğin, oksin, sonradan doğrudan uyarıcı etki gösterebilen başka bir hormonun (etilen) üretiminde artış sağlayarak dolaylı etki gösterebilir.
Öksinler (bu anlamda genel olarak köklendirme hormonları olarak da adlandırılırlar), gövde ya da yaprak gibi organların çeliklerinden adventif köklerin geliştirilmesinde önemli bir ticari kullanım bulmuşlardır. Sardunyalar ve sögütler gibi bazı bitkilerin çelikleri, hormon uygulanmaksızın, suda ya da toprakta kolayca köklenmelerine karşın, bitkilerin çoğu bu şekilde çoğaltılamaz. Böyle durumlarda oksin uygulanması, genellikle kök oluşumunu teşvik etmektedir. Böylelikle, kaybedilme olasılığı olan pek çok değerli genotipin vejetatif olarak çoğaltılması mümkün olmaktadır.
Kimyasal Yabani Ot Kontrolü![Oksin Hormonunun Görevleri]()

2,4-diklorofenoksiasetik asit, yaygın olarak kullanılan modern bir ot öldürücü ya da herbisittir. Genellikle 2,4-D olarak kısaltılır. Oksin testlerinin tümüne yanıt vermesede sentetik kimyasal, oksinin pek çok özelliğini gösterir. Oksin benzeri sentetik kimyasallar, 1940’lardan bu yana, karahindiba benzeri yapraklı türlerin ve diğer geniş yapraklı türlerin kontrolünde bol miktarda kullanılmışlardır. Uygun konsantrasyonlarda kullanıldıklarında, seçicilik göstermeleri ve çimler ile bunları n akrabaları olan diğer monokotilleri öldürmemeleri nedeniyle, çimenliklerde, otlaklarda, mısır, buğday, çavdar ve pirinç tarlalarında geniş yapraklı türlerle mücadelede çok büyük ticari öneme sahiptirler. Bu sentetiklerin düşük konsantrasyonlarda doğal oksinlerle benzer etkiler göstermelerine karşın, yüksek konsantrasyonlarda bazı bitki kısımlarında büyümeyi engelleyip, diğer kısımlarında ise hızlı, koordinasyonsuz ve düzensiz bir büyümeyi teşvik ederek bitkileri öldürürler. Ne bu sonuçların tam oluşma şekli ne de çimlere göre (monokotillev) geniş yapraklı türlerin daha duyarlı olmalarının nedeni bilinmemektedir.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç