
 Canlı organizmaların populasyonları sınırsız bir şekilde büyümezler; bunların boyutlarını sınırlayan bir ya da daha fazla faktör mutlaka vardır. Populasyonların çoğu, onları en iyi karakterize eden büyüme eğrilerine göre adlandırılır. Örneğin, bu kavramsal skalanın bir ucunda yüksek rmax’a sahip çok hızlı üreyen populasyonlar bulunur. Bunlar bu kapasiteyi geçiş olabilecek, bozulmuş ya da yeni çevrelerde kullanabilmek üzere, geliştirmişlerdir. Sivrisineklerin ilkbaharda hızla çoğalması, bir rmax türünün davranışını örnekler. Neyse ki, bizim için sivrisinek populasyon büyüklüğü, genel kuraklık ve soğuk gibi çevre faktörleri tarafından baskı altında tutulmaktadır.
Canlı organizmaların populasyonları sınırsız bir şekilde büyümezler; bunların boyutlarını sınırlayan bir ya da daha fazla faktör mutlaka vardır. Populasyonların çoğu, onları en iyi karakterize eden büyüme eğrilerine göre adlandırılır. Örneğin, bu kavramsal skalanın bir ucunda yüksek rmax’a sahip çok hızlı üreyen populasyonlar bulunur. Bunlar bu kapasiteyi geçiş olabilecek, bozulmuş ya da yeni çevrelerde kullanabilmek üzere, geliştirmişlerdir. Sivrisineklerin ilkbaharda hızla çoğalması, bir rmax türünün davranışını örnekler. Neyse ki, bizim için sivrisinek populasyon büyüklüğü, genel kuraklık ve soğuk gibi çevre faktörleri tarafından baskı altında tutulmaktadır.
Yüksek rmax’lu yaşam biçimi, genellikle bir dili davranış ve fizyolojik adaptasyonla birlikte görülür. Örneğin, bireyler, çok sayıda küçük ve hızla olgunlaşan yavrular oluştururlar. Çünkü şiddetli rekabet olmadığında, böyle bireylerin uyumu, bazı çevresel etkiler ani bir azalmaya yol açmadan önce, mümkün olabildiği kadar çok sayıda genç birey üretmeye bağlıdır. Böyle bir türün hayatta kalma eğrisi genellikle III. tipteki gibidir.
Bu kavramsal skalanın diğer ucunda; ortamın taşıma gücündeki (K). ya da ona yakın olan populasyonlar bulunur, bunlar çok yavaş ürerler ya da hiç çoğalmazlar. Filler gibi büyük memelilerin çoğunun örnek oluşturduğu bu türlerde, düşük rmax uygun hale getirilmiştir. Düşük rmax türleri tipik olarak, kararlı ya da bilinen ortamlarda bulunurlar.
Bunların uyumları, hızlı üremeden ziyade, sınırlı kaynaklar için etkili rekabet edebilme yeteneklerine bağımlı olduğundan, böyle türlerin bireyleri, sadece birkaç tane, uzun ömürlü, yavaş erginleşen gençler üretir; nicelikten ziyade nitelik önemlidir.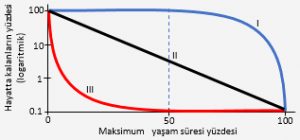
Bu türlerin hayatta kalma eğrileri I. tipe benzer. Rmax spektrumunun bu iki ucu, içinde çeşitli populasyon kontrol mekanizmalarının işlediği yolları, çarpıcı alarak açıklamaktadır. Görmüş olduğumuz gibi, yüksek rmax türlerinde populasyon yoğunluğu genellikle, besin, su, yuvalanma alanı gibi gerekli kaynakların uygunluğu ile sınırlanmak için çok düşüktür. Bunlar şiddetli bir predasyona maruz kalsalar bile tekrar çok hızla ve bol olarak ürerler ve bu faktör pek sınırlayıcı olmayabilir. Populasyonu periyodik olarak azaltan hava, yangın, sel gibi yoğunluğa bağlı olmayan faktörler, daha önemli bir yer tutar. Diğer yandan, düşük rmax türlerin populasyonlarının, bir ortamın taşıma kapasitesinin çok üzerinde büyümesi bir ya da birden fazla kritik kaynağın sınırlı oluşu gibi faktörler tarafından engellenir. Yani bu türler hem yoğunluğa bağlı, hem de yoğunluğa bağlı olmayan faktörlerden etkilenir.
Yoğunluğa bağlı sınırlayıcı faktörler olarak, predasyon, parazitizm ve hastalıklar
Predasyon, hastalık ve parazitizm genellikle, avı (veya konakçıyı) yoğunluğa bağlı olarak etkiler. Konakçı ya da av popülasyonu arttıkça, populasyonun büyük bir yüzdesi zarar görür; çünkü, bireyler uygun olmayan koşullar içinde bulunmaya zorlanarak ya da mevcut kaynaklardan daha az yararlanılması nedeniyle zayıf düştüklerinden daha kolay bulunabilir ve saldırıya uğrar. Aynı şekilde, bir parazitin, uygun bir konakçı bulma olası lığı da, genellikle yoğunluğa bağlıdır. Bir dizi av türünü yakalayan parçalayıcılar avlanma biçimlerini, aylarının nisbi yoğunluklarına göre, daha yaygın türlere konsantre olacak şekilde değiştirme eğilimine girerler. Eğer av olan türün yoğunluğu artarsa, onun üzerinden beslenen predatörlerin yoğunluğu da artar. Belirli av türleri üzerinde konsantrasyonlarının artması ile birlikte predatörlerdeki bu artış, av yoğunluğunun tekrar düşüşüne neden olan bir faktör olabilir. Ancak, ayın yoğunluğu azaldıkça, genellikle predatör türlerin yoğunluğunda da biraz gecikmiş bir azalma görülür. Sonuç, seri popülasyon değişimleri olabilir, predatör ve av populasyonlarındaki bu bağlantılı değişimler, predatör için en önemli sınırlayıcı faktörün besinin bulunabilirliği olduğunu ve bu nedenle predasyonun av için, önemli bir sınırlayıcı faktör olduğunu göstermektedir.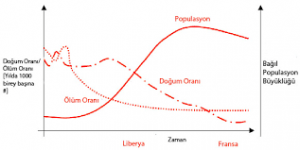
Predatör potansiyel av olabilecek türlerden en yaygın olanına yönelebildiğinde ya da ortamsal heterojenite özelleşmiş türler üzerinde aylanan nadir bireyler için bir sığınak sağladığında, hem predatör, hem de ayın kararlı populasyonları varlıklarını sürdürebilir. Fakat, ilişkilerin çoğunda populasyon döngüleri kuraldır.
Predatör-av dengesi, iyi niyetli kontrol yöntemleri ile kasıtsız olarak bozulabilir. Örneğin, meyvelere zarar veren siklamen akarlarını yok etmek için çileklere belirli insektisitlerin uygulanması, hem siklamen akarlarını, hem de onlarla beslenen karnivor akarları öldürmüştür.
Fakat yüksek rmax’lu ve predasyon’dan kurtulmuş siklamen akarları , çilek tarlalarını tekrar hızlı bir şeklide sararken, predatör akarlar çok daha yavaş geri dönerler. Sonuçta, doğal predatörleri olmayan siklamen akarlarının yoğunlukları hızla artmış ve hiç bir zaman insektisit uygulanmamış duruma göre çok daha fazla zarar vermişlerdir.
Bu, av (zararlı) eklembacaklılara göre, predatör (faydalı) eklembacaklıların daha yavaş toparlanmaları çok yaygındır; bunun ötesinde zararlılar için daha yüksek rmax büyük bir olasılıkla, onların predatörlerinden daha önce pestisitlere direnç kazanmış oldukları anlamına gelir. Bu faktörler, tarımda aşırı pestisit kullanımının uzun dönemdeki yararı üzerinde kuşku yaratmaktadır.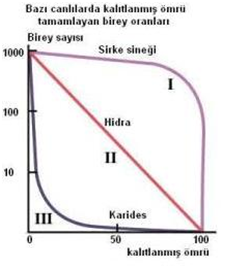
Tahmin edileceği gibi, predatörlerin davranışı, av popülasyonlarının üreme stratejilerini büyük ölçüde etkileyebilir. Örneğin, bazı yörelerde yaşayan lepistesler, erginler üzerinde özelleşmiş büyük predatörlere maruz kalırken, diğer habitatlarda, tehdit, gençler üzerinde avlanan küçük balıklardan gelir. Ergin-predatör habitatındaki le-pistesler, genç evre-predatör habitatından yaklaşık iki kat fazla yavru üretirler. İlk gruptaki yavrular, diğerlerinin yaklaşık yarısı kadar ağırlıktadır. Bir rmax türünde bu modifikasyonlar, aynı predatörleri kullanarak ve 30-60 jenerasyon bekleyerek, laboratuvarda da yaratılabilir.
Hastalıklar, çeşitli yollarla populasyon büyüklüğünü sınırlarlar. Patojenler duyarlı bireyler üzerinde sabit bir baskı yaratırlar; ancak, kırmızı kraliçe seçilim hipotezinde, konakçı türler (seleksiyonun bağışıklığı uygun hale getirdiği) ve parazitler (seleksiyonun hedef populasyon hücrelerini daha iyi tanıyabilme şansı verdiği) arasında değişmez bir yarış vardır. Bağışıklık, konakçı bireylerin sağlığına da bağlı olduğundan, hastalıklar zayıf üyeleri gruptan atarak, populasyonu habitatlarının halihazırdaki taşıma kapasitelerinin üzerinde sınırlamak için etki edebilir. Yüksek populasyon yoğunluğu birçok hastalığın geçişini de kolaylaştırır.
Sonuç olarak, hastalıklar, AIDS virüsünün bir klonunun daha önceki primat konakçısının (muhtemelen Afrika yeşil maymunları ) yanında insanlara da bulaşmasını sağlayan bir mutasyonu kazanmasında olduğu gibi patojenler yeni konakçı türleri enfekte ettiklerinde populasyon büyüklüğünü değiştiren önemli bir şans elde ederler.
Konakçı yayılımının genişlemesine ilişkin daha çarpıcı bir örnek, bir köpek distemper virüsünün kuzey batı Avrupa kıyılarında, dört ay içinde 12.000’den fazla ölüme yol açarak fokları infekte etmeye başladığı 1988 yılında ortaya çıkmıştır. Hastalıklar daha önceden mevcut coğrafik engeller ortadan kalktığında da önemli bir etki yaratabilir.
Bu durum patojenin seçilimle bağışıklık kazanmamış popülasyonlara ulaşmasına izin verir. Avrupalıların onbeşinci ve onaltıncı yüzyılda Amerika’ya ulaştıklarında, çiçek gibi yeni hastalıklar taşıyarak, yerlilerde kitle ölümlerine neden olması bu evrimsel olgu ile açıklanabilir.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç