
Bir impulsun bir sinir hücresinin aksonu boyunca nasıl iletildiğini anlamak, nöronların kendi aralarında nasıl haberleştiğini ve sonuçta sinirsel kontrolün nasıl başarıldığını anlamada ilk adımdır. Bir nöronun aksonu genelde diğer nöronların dendritleri ya da hücre gövdesiyle sinaps yapar. Bir aksonun son ucu normalde çok sayıda dallanma yaptığı için bir tek nöron birçok başka nöronla sinaps yapabilir. Hatta bunun da ötesinde genellikle bu nöronların her biriyle çok sayıda noktada sinaps yapar. Bir aksonun ince dallarından her biri, düğme şeklinde küçük şişkinliklerle sona erer. Bu yapılara sinaptik uç denir. Az sayıda olmak üzere bazen sinaptik uç zarı ile bununla temasta olan hücrenin zarı arasında bir iletim bölgesi (gap junction) bulunur. Böyle bir bölge, iki nöron arasında doğrudan bir 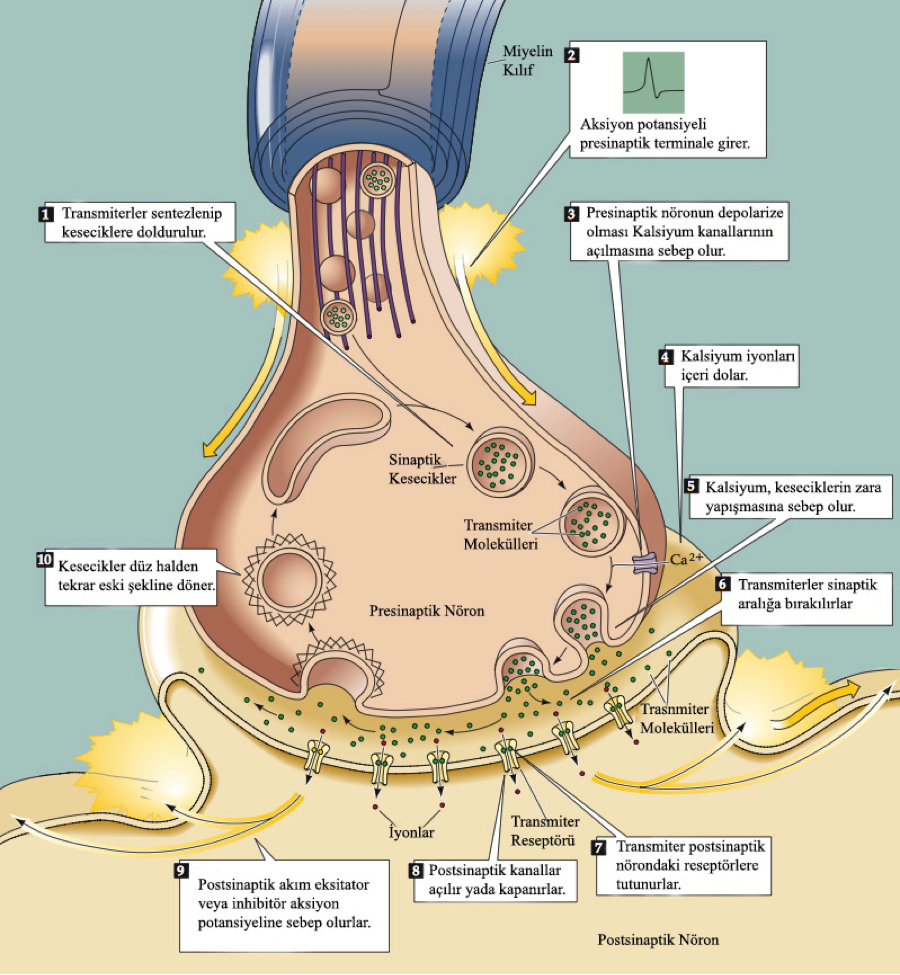 elektriksel iletim sağlar ve böylece birinci nöronun aksonu boyunca ilerleyen impuls, hemen hiçbir dirençle karşılaşmadan, ikinci nörona geçer. Elektriksel sinapslar impuls iletimindeki gecikmeyi en aza indirdikleri için, sinir sisteminde iletim hızının özellikle önemli olduğu yerlerde görülürler. Bunlar ayrıca, birinci nörondaki impulsun ikinci nöronda da bir impuls yaratmasını büyük ölçüde güvenceye alır.
elektriksel iletim sağlar ve böylece birinci nöronun aksonu boyunca ilerleyen impuls, hemen hiçbir dirençle karşılaşmadan, ikinci nörona geçer. Elektriksel sinapslar impuls iletimindeki gecikmeyi en aza indirdikleri için, sinir sisteminde iletim hızının özellikle önemli olduğu yerlerde görülürler. Bunlar ayrıca, birinci nörondaki impulsun ikinci nöronda da bir impuls yaratmasını büyük ölçüde güvenceye alır.
Buna karşın sinapsların çok büyük bir çoğunluğu elektriksel değil kimyasaldır. Birinci nöronun (presinaptik) sinaptik ucu ile ikinci nöronun (postsinaptik) zarını ayıran, yaklaşık 20 nm genişliğindeki boşluğa sinaptik aralık adı verilir. Bu boşluk boyunca iletim son uçta yer alan küçük sinaptik veziküllerden salınıp difüze olan transmitter kimyasallar tarafından gerekleştirilir. Bu binlerce vezikülden herbirinde 10.000 taneye kadar transmitter molekülü bulunur.
Presinaptik nöronun aksonu boyunca ilerleyen impuls terminale ulaşınca sinaps bölgesinde yoğunlaşmış olan özel voltaj-kapılı kalsiyum kanalları açılır ve son uçtaki zar Ca++ iyonlarına daha geçirgen hale gelir. Hücre dışında 10.000 kat daha yoğun olarak bulunduklarından, Ca++ iyonlarını n bir şekilde uyarmasıyla son uçtaki sinaptik veziküller, ucun zarına doğru ilerleyip bununla kaynaşırlar ve parçalanırlar. Böylece transmitter kimyasallar egzositoz yoluyla sinaptik boşluğa şalınırlar. Ca++ iyonları kas kasılmasını başlatmak için de benzer bir rol oynarlar.
Sinaptik boşluğa salınan transmitter molekülleri, difüzyonla ilerleyip ikinci nöronun postsinaptik zarında yer alan yüksek derecede özgül reseptörlere zayıf bağlarla bağlanırlar. Bu reseptörler nörotransmittere özgüldür ve hormon reseptörlerininkine benzer biçimde işlev görürler.
Transmitter asetilkolin (omurgalı motor nöronlarının kas hücreleriyle iletişim kurmasını sağlayan kimyasal) olduğunda, reseptörü aktive etmek için iki transmitter molekülünün buna bağlanması gerekir. Bir tek vezikülün içerdiği moleküller yaklaşık 2000 reseptörü aktive eder. Transmitterin reseptöre bağlanması, bir kanalın kapılarını açar ve özgül bir iyonun zarı geçmesini sağlar. Bu iyon hareketi postsinaptik nöronun zarında, potansiyel değişikliğiyle sonuçlanır ve o hücrede yeni bir impuls yaratılabilir. Asetilkolin reseptörlerini taşıyan kanallar hem K+ hem de Na+ iyonlarını geçirirler ve hücrenin bu kanalların etrafındaki bölgeleri kısmen depolarize olur. Bir sinapsta iletim, bir dizi olay gerektirdiğinden -son uca Ca++ iyonlarını girmesi, transmitter veziküllerinin hareketi, egzositoz, transmitterin sinaptik boşlukta difüzyon ve en sonunda iyonların postsinaptik kanallar içinde difüzyonu- işlem, impulsun nöron boyunca iletilmesinden çok daha yavaştır. Bu nedenle, bir mesajın iletilme süresi beklenenden uzun olacaktır. Genel olarak, bir sinir yolunda ne kadar çok sinaps varsa, yol boyunca birim uzunluk başına ortalama iletim hızı o kadar yavaştır.
Transmitterin sinaptik boşlukta difüze olup impulsun postsinaptik hücreye aktarılması, olayın sonu değildir. Eğer transmitter kalıcı olursa postsinaptik reseptörler, bir tek impulsun gelmesiyle sonsuza kadar uyarılırlar. O zaman, transmitteri ortadan kaldıracak bir mekanizma olmalıdır. Örneğin, asetilkolin, sinaptik boşluktan difüze olup postsinaptik hücrenin dendriti ya da hücre gövdesi zarı üzerinde etkisini gösterdikten sonra, asetilkolinesteraz adı verilen bir enzim tarafından hızla inaktive edilir. Bu enzim, transmitteri ortadan kaldırmakla, yeni bilgi taşıyan bir sonraki impulsun iletilmesine olanak sağlar. Birçok insektisit, örneğin organofosfatlar (sinir gazları olarak da bilinirler) kolinesteraz 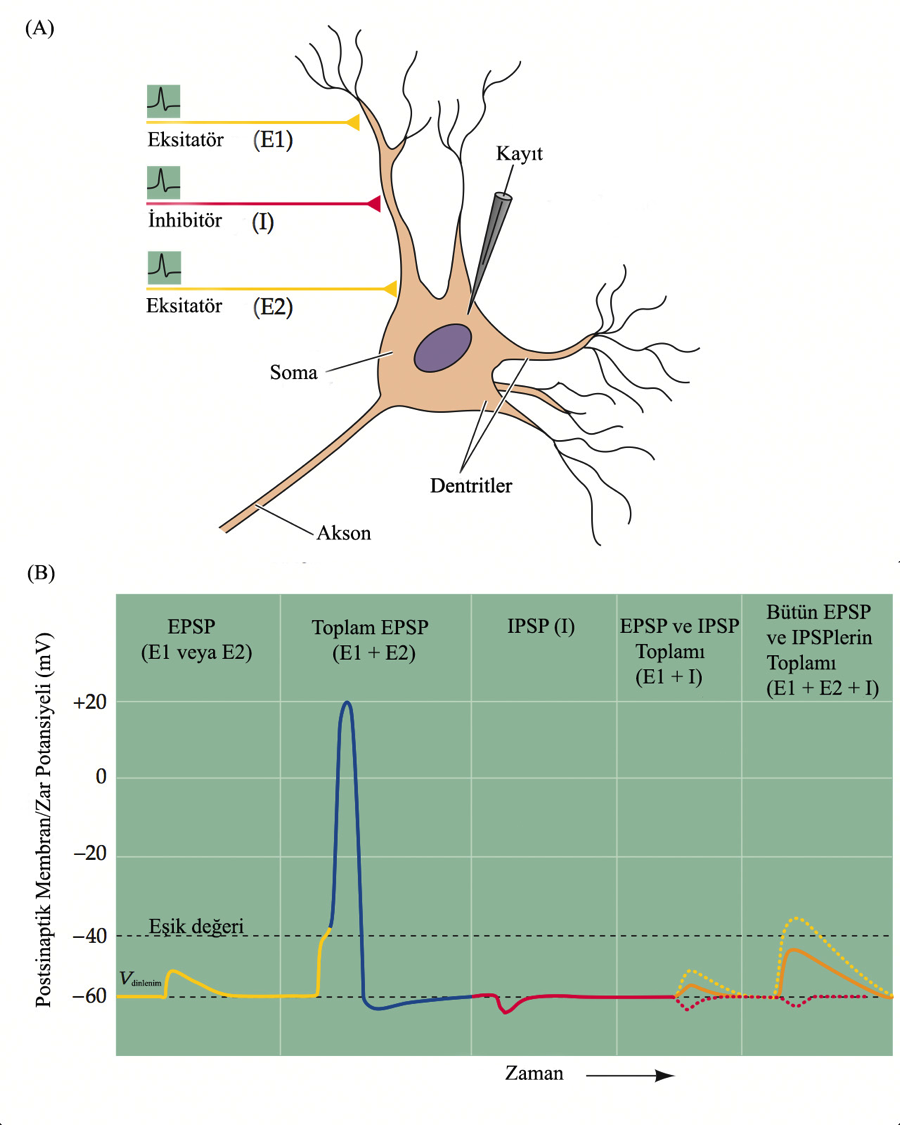 inhibitörüdür. Bunlar asetilkolinin ortadan kaldırılmasını önlerler ve bunun sonucunda, beklendiği gibi, bunlara maruz kalan bir böceğin sinapsları sürekli olarak aktif kalır. Yeteri kadar yüksek dozda verilecek olurlarsa kolinesteraz inhibitörleri önemli fizyolojik olayları etkilerler ve hayvan ölür.
inhibitörüdür. Bunlar asetilkolinin ortadan kaldırılmasını önlerler ve bunun sonucunda, beklendiği gibi, bunlara maruz kalan bir böceğin sinapsları sürekli olarak aktif kalır. Yeteri kadar yüksek dozda verilecek olurlarsa kolinesteraz inhibitörleri önemli fizyolojik olayları etkilerler ve hayvan ölür.
Merkezi sinir sistemi dışında bir transmitter olarak işlev gören asetilkolin, merkezi sinir sisteminde de bulunan ve giderek büyüyen transmitterler listesinde de yer alır. Omurgalılarda bunlar arasında noradrenalin (aynı zamanda adrenal medullada da bir hormon olarak yapılır), serotonin, dopamin, azot oksit ve gamma amino-butirik asit (GABA) bulunur. Daha önceleri nedeni belirsiz ruhsal bozukluklardan kaynaklandıkları sanılan şizofreni ve ciddi depresyon gibi çeşitli anormalliklerin, şimdi artık MSS transmitterlerinin, reseptörlerin ve daha önce bilinmeyen nöral hormonların biyokimyasal aksaklıklarından ileri geldiği bilinmektedir. Bu keşifler bazı ruhsal bozuklukların nispeten kesin fizyolojik tedavileri için yol açmaya başlamışlardır
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç