
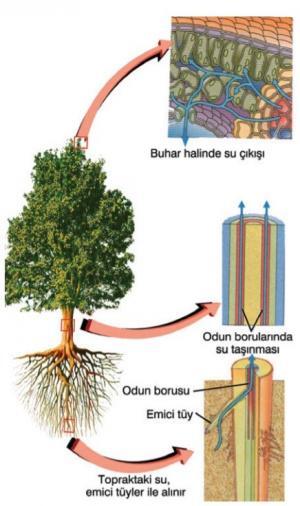
 Yağmur suyu, bitkilere, kapilar su olarak isimlendirilen ve toprak partiküllerini çevreleyen gevşek bir su filmi olarak yayılmıştır. Kökler, özellikle de kök tüyleri, bu suyla temas halindedirler. Kökteki epidermisin kutikulası bulunmadığından, kapiler su, köklere kolaylıkla iki şekilde girebilir. Su, basit olarak, bir kök tüyünün zarını geçtikten sonra korteks içinde hücreden hücreye taşınabilir ve endodermisi geçerek kökün merkezi silindirine ulaşabilir. Hücreler arasındaki bu taşınım, kök tüyünün zarı geçildikten sonra diğer zarlar tarafından engellenmez. Çünkü, hatırlanacağı gibi, birbirine komşu bitki hücrelerinin stoplazmaları genellikle bir simplast oluştururlar. Simplast, plazmodezmata ile hücrelerin içeriklerinin birbirine bağlanmasıyla oluşan bir topluluktur. Suyun diğer giriş yolu apoplasttır. Apoplast, korteks’e kadar uzanan, hücre çeperleri ve hücreler arası boşlukların oluşturduğu bir ağdır. Kaspari şeridi suyun apoplastta daha fazla ilerlemesini önler. Bu nedenle suyun endodermis tabakası ndaki bir hücrenin zarından geçmesi gerekir. Suyun, kökün öz bölgesine simplast ya da apoplast ile ulaşmasına bakılmaksızın, bir bitki hücresinin zarını geçebilmesi için bazı noktalarda teşvik edilmesi gerekir. Bunun, basit olarak, ozmozis ile nasıl gerçekleştiğini görebiliriz: su, eriyik konsantrasyonu düşük olan bölgelerden, eriyik konsantrasyonu yüksek olan bölgelere akar. Sitoplazma çok yüksek konsantrasyonlarda iyonlar, şekerler ve diğer hidrofilik organik moleküllerden oluşan çözünmüş maddeleri içerir. Bu nedenle, su, köklere girme eğilimindedir.
Yağmur suyu, bitkilere, kapilar su olarak isimlendirilen ve toprak partiküllerini çevreleyen gevşek bir su filmi olarak yayılmıştır. Kökler, özellikle de kök tüyleri, bu suyla temas halindedirler. Kökteki epidermisin kutikulası bulunmadığından, kapiler su, köklere kolaylıkla iki şekilde girebilir. Su, basit olarak, bir kök tüyünün zarını geçtikten sonra korteks içinde hücreden hücreye taşınabilir ve endodermisi geçerek kökün merkezi silindirine ulaşabilir. Hücreler arasındaki bu taşınım, kök tüyünün zarı geçildikten sonra diğer zarlar tarafından engellenmez. Çünkü, hatırlanacağı gibi, birbirine komşu bitki hücrelerinin stoplazmaları genellikle bir simplast oluştururlar. Simplast, plazmodezmata ile hücrelerin içeriklerinin birbirine bağlanmasıyla oluşan bir topluluktur. Suyun diğer giriş yolu apoplasttır. Apoplast, korteks’e kadar uzanan, hücre çeperleri ve hücreler arası boşlukların oluşturduğu bir ağdır. Kaspari şeridi suyun apoplastta daha fazla ilerlemesini önler. Bu nedenle suyun endodermis tabakası ndaki bir hücrenin zarından geçmesi gerekir. Suyun, kökün öz bölgesine simplast ya da apoplast ile ulaşmasına bakılmaksızın, bir bitki hücresinin zarını geçebilmesi için bazı noktalarda teşvik edilmesi gerekir. Bunun, basit olarak, ozmozis ile nasıl gerçekleştiğini görebiliriz: su, eriyik konsantrasyonu düşük olan bölgelerden, eriyik konsantrasyonu yüksek olan bölgelere akar. Sitoplazma çok yüksek konsantrasyonlarda iyonlar, şekerler ve diğer hidrofilik organik moleküllerden oluşan çözünmüş maddeleri içerir. Bu nedenle, su, köklere girme eğilimindedir.
 Bitki bilimcileri, suyun hareketine su potansiyeli açısından bakmayı yeğlerler, yani, zar’ın karşı lı klı iki tarafında suyun potansiyel enerjisini karşılaştı rırlar. Bu açıdan, içerdiği eriyik miktarı yüksek olan suyun potansiyeldi, daha fazla eriyik içeren sudan daha yüksektir. Dolayısıyla su, yüksek potansiyeli olan bölgelerden düşük potansiyelli bölgelere akar. Suyun hareketi ile ilgili bu bakış açısı, nispi ozmotik konsantrasyonun dışındaki etmenlerin çoğunlukla önemli olması nedeniyle, özellikle bitkiler için uygundur. Özelde, bitkiler sert bir hücre çeperine sahip olduklarından, hücredeki basınç (turgor basıncı olarak adlandırılır) sıfırlık bir su potansiyeli üretecek şekilde, ozmotik basınç farkını tam dengeleyen bir potansiyel enerji üretmeden önce yalnızca sınırlı miktarda su alırlar. İleride görüleceği gibi, köklerden net bir su girişinin sürdürülmesi bitkiler açısından, önemli olduğundan, köklerde sıfır ya da negatif su potansiyeli oluşumunun önlenmesine yardımcı olan mekanizmalar geliştirilmiştir. Suyun, epidermis hücresine girdikten sonra, gerekli olduğu yerlere, genellikle de bitkinin yapraklarına ve büyümekte olan kısımlarına ulaştırılması gerekir. Bitkiler, her zaman kökün dış kısmından ksileme yöneldikçe daha düşük bir su potansiyeline sahip olduklarından bu yavaş, ancak otomatik olarak ortaya çıkar. Bu potansiyel iki şekilde oluşturulur. Suyun yaprak hücrelerinden buharlaşmayla kaybı, yapraklardaki su potansiyelinin çok düşüren baskın etmendir; başka bir deyişle, yapraklara turgor basıncı düşmüş, ozmotik konsantrasyon çok artmıştır. Bunun sonucunda, suyu köklerden aynı şekilde alınmasına sağlayan, iletim sistemindeki su potansiyeli düşürülerek su, ksilemden yukarı çekilir. Nadiren evaporasyon hızı düşük olduğunda (örneğin gökyüzü bulutlu ya da karanlık iken hava soğuk ve nemli olduğunda), moleküler pompalar iyonları ksileme taşıyarak onun su potansiyelini düşürürler (önemli bir metabolik maliyeti olsa bile). Bu strateji, toprak nispeten kuru ve dolayısıyla düşük bir su potansiyeline sahip olduğu durumlarda da yardımcı olabilir.
Bitki bilimcileri, suyun hareketine su potansiyeli açısından bakmayı yeğlerler, yani, zar’ın karşı lı klı iki tarafında suyun potansiyel enerjisini karşılaştı rırlar. Bu açıdan, içerdiği eriyik miktarı yüksek olan suyun potansiyeldi, daha fazla eriyik içeren sudan daha yüksektir. Dolayısıyla su, yüksek potansiyeli olan bölgelerden düşük potansiyelli bölgelere akar. Suyun hareketi ile ilgili bu bakış açısı, nispi ozmotik konsantrasyonun dışındaki etmenlerin çoğunlukla önemli olması nedeniyle, özellikle bitkiler için uygundur. Özelde, bitkiler sert bir hücre çeperine sahip olduklarından, hücredeki basınç (turgor basıncı olarak adlandırılır) sıfırlık bir su potansiyeli üretecek şekilde, ozmotik basınç farkını tam dengeleyen bir potansiyel enerji üretmeden önce yalnızca sınırlı miktarda su alırlar. İleride görüleceği gibi, köklerden net bir su girişinin sürdürülmesi bitkiler açısından, önemli olduğundan, köklerde sıfır ya da negatif su potansiyeli oluşumunun önlenmesine yardımcı olan mekanizmalar geliştirilmiştir. Suyun, epidermis hücresine girdikten sonra, gerekli olduğu yerlere, genellikle de bitkinin yapraklarına ve büyümekte olan kısımlarına ulaştırılması gerekir. Bitkiler, her zaman kökün dış kısmından ksileme yöneldikçe daha düşük bir su potansiyeline sahip olduklarından bu yavaş, ancak otomatik olarak ortaya çıkar. Bu potansiyel iki şekilde oluşturulur. Suyun yaprak hücrelerinden buharlaşmayla kaybı, yapraklardaki su potansiyelinin çok düşüren baskın etmendir; başka bir deyişle, yapraklara turgor basıncı düşmüş, ozmotik konsantrasyon çok artmıştır. Bunun sonucunda, suyu köklerden aynı şekilde alınmasına sağlayan, iletim sistemindeki su potansiyeli düşürülerek su, ksilemden yukarı çekilir. Nadiren evaporasyon hızı düşük olduğunda (örneğin gökyüzü bulutlu ya da karanlık iken hava soğuk ve nemli olduğunda), moleküler pompalar iyonları ksileme taşıyarak onun su potansiyelini düşürürler (önemli bir metabolik maliyeti olsa bile). Bu strateji, toprak nispeten kuru ve dolayısıyla düşük bir su potansiyeline sahip olduğu durumlarda da yardımcı olabilir.
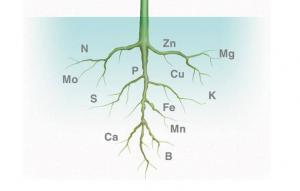
 Bitkilerin, yapraklardan köklere, uygun bir su potansiyeli gradiyentini nasıl koruduklarını görmüş bulunuyoruz; şimdi besin elementlerini su ile nasıl alındıklarını ve köklerin besin elementi alınımını nasıl düzenlediklerini görelim. Bitkiler mineralleri, genellikle iyonik yapıda alırlar: azot; nitrat (NO-3) ya da amonyum (NH+4) iyonları; fosfor; dihidrojen fosfat (H2P0+4) ya da monodihrojen fosfat (HP0-4) iyonları; kükürt; sülfat iyonları (S0+4); potasyum, kalsiyum, magnezyum ve demir ise bu minerallerin basit iyonları (K+, Ca++, Mg++ ve Fe++ ya da Fe+++) şeklinde absorbe edilir. Bitkilerin absorbe edebilecekleri iyonlar toprak suyu çözeltisinde bulunurlar. Bunların konsantrasyonları, toprağın verimliliğine, asitliliğine ve diğer etmenlere bağlı olarak değişiklik gösterir. Toprak mineralleri, çözeltide bulunmayı’) iyonik bağlarla toprak partiküllerine bağlı olduklarında bitkiler için yararlı değillerdir. Bağlı olan bu minerallerin daha serbest hale geçirilerek kökler tarafından absorbe edilebilmesini sağlamak için, tarım yapılan toprakların asitliliği sıklıkla değiştirilir. Örneğin, pH’ı n yükseltmek için çok asidik toprağa kireç ilavesi, fosfor, potasyum ve molibdenin yarayışlılığını arttırabilir; ancak aşırı kireç, yarayışlı demir, bakır, mangan ve çinkoyu azaltabilir.
Bitkilerin, yapraklardan köklere, uygun bir su potansiyeli gradiyentini nasıl koruduklarını görmüş bulunuyoruz; şimdi besin elementlerini su ile nasıl alındıklarını ve köklerin besin elementi alınımını nasıl düzenlediklerini görelim. Bitkiler mineralleri, genellikle iyonik yapıda alırlar: azot; nitrat (NO-3) ya da amonyum (NH+4) iyonları; fosfor; dihidrojen fosfat (H2P0+4) ya da monodihrojen fosfat (HP0-4) iyonları; kükürt; sülfat iyonları (S0+4); potasyum, kalsiyum, magnezyum ve demir ise bu minerallerin basit iyonları (K+, Ca++, Mg++ ve Fe++ ya da Fe+++) şeklinde absorbe edilir. Bitkilerin absorbe edebilecekleri iyonlar toprak suyu çözeltisinde bulunurlar. Bunların konsantrasyonları, toprağın verimliliğine, asitliliğine ve diğer etmenlere bağlı olarak değişiklik gösterir. Toprak mineralleri, çözeltide bulunmayı’) iyonik bağlarla toprak partiküllerine bağlı olduklarında bitkiler için yararlı değillerdir. Bağlı olan bu minerallerin daha serbest hale geçirilerek kökler tarafından absorbe edilebilmesini sağlamak için, tarım yapılan toprakların asitliliği sıklıkla değiştirilir. Örneğin, pH’ı n yükseltmek için çok asidik toprağa kireç ilavesi, fosfor, potasyum ve molibdenin yarayışlılığını arttırabilir; ancak aşırı kireç, yarayışlı demir, bakır, mangan ve çinkoyu azaltabilir.
Her bir mineralin kökler tarafından absorbsiyon hızı, suyun ve diğer minerallerin absorbsiyon hızlarından büyük ölçüde bağımsızdır. Her bir besleyici, onun kök içindeki ve dışındaki konsantrasyonu, iyon kanallarından hücre zarlarına pasif olarak geçebilme kolaylığı ve taşıyıcı molekül miktarı gibi etmenlerin belirlediği bir hızda köke girer. Konsantrasyon gradienti bitkinin içine girişi kolaylaştırdığında, absorbsiyon hızı, tek başına pasif difüzyonla alınandan daha büyüktür. Burada, kolaylaştırılmış bir difüzyon söz konusudur. Bununla birlikte, bitkiler, sı klı kla kök hücreleri içindeki konsantrasyonları toprak çözeltisindeki konsantrasyonlardan daha fazla olan mineralleri alırlar. Eğer bu alma işlemi yalnızca basit difüzyonla gerçekleşseydi bunun olmaması gerekirdi. Burada aktif bir taşınımın rölü olduğu açıktır; bitki, varlığını sürdürebilmek için gerekli mineral besleyicilerin alınmasında enerji harcar. Bu kitabın tümünde görüleceği gibi, bitki ya da hayvan olsun, maddeler, canlı hücrelerin zarlarında madde geçişinde aktif taşınım organizmaların çoğunda bir istisna değil, genel bir kural olarak ortaya çıkar.
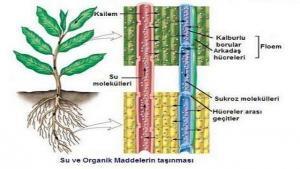 Kök hücrelerinde minerallerin yüksek konsantrasyonlarda birikmelerini önleyen iki etmen, bu maddelerin en hızlı şekilde köklere girişine yardımcı olur: Bir mineral köke girdikten sonra oradan uzaklaştırılarak bitkinin diğer bazı kısımlarına taşınabilir; ayrıca, hızla farklı bir bileşiğin sentezinde kullanılabilir. Örneğin, nitrat halinde absorbe edilen azot hızla indirgenerek amino asitler ve amidler gibi azotça zengin bileşiklerin yapımında kullanılır. Bu bileşikler daha sonra taşını rlar ve biriktiririlirler. Birikimin çoğu kofullarda gerçekleşir. Burada azotlu bileşiklerin konsantrasyonu, genellikle sitoplazmanınkinden daha fazladır. Bu, koful zarı ya da tonoplastın, bileşiklerin vakuol’e girişine izin verdiklerini, ancak oradan çıkışlarını önleyerek seçicilik gösterdiklerini açıkça göstermektedir.
Kök hücrelerinde minerallerin yüksek konsantrasyonlarda birikmelerini önleyen iki etmen, bu maddelerin en hızlı şekilde köklere girişine yardımcı olur: Bir mineral köke girdikten sonra oradan uzaklaştırılarak bitkinin diğer bazı kısımlarına taşınabilir; ayrıca, hızla farklı bir bileşiğin sentezinde kullanılabilir. Örneğin, nitrat halinde absorbe edilen azot hızla indirgenerek amino asitler ve amidler gibi azotça zengin bileşiklerin yapımında kullanılır. Bu bileşikler daha sonra taşını rlar ve biriktiririlirler. Birikimin çoğu kofullarda gerçekleşir. Burada azotlu bileşiklerin konsantrasyonu, genellikle sitoplazmanınkinden daha fazladır. Bu, koful zarı ya da tonoplastın, bileşiklerin vakuol’e girişine izin verdiklerini, ancak oradan çıkışlarını önleyerek seçicilik gösterdiklerini açıkça göstermektedir.
Kök sistemleri yeterli olsa bile, dışardan yardım almaksızın, bitkilerin yaşamlarını sürdüremeyecekleri birçok toprak bulunmaktadır. Bunun nedeni ya uygun besin elementlerinin bulunmayışı ya da besin elementlerinin bitkiler tarafından alınabilecek yapıda olmamalarıdır. Bununla birlikte, daha önce belirtmiş olduğumuz gibi, bazen mantarlar ve bitki kökleri mikoriza olarak bilinen birlikler oluştururlar. Bunlar bitkilerin mineral alınımını büyük ölçüde kolaylaştırabilirler. Bazıları toprak yüzeyinde şapkalı mantarlar olarak ortaya çıkan mantarlar, aslında konukçullarının kök hücrelerine girerek besleyicileri doğrudan içeriye alırlar. Bu nedenle, bu ilişki, karşılıklı olarak avantaj sağlayıcı ya da mutualistiktir.
Kaynakça:
Biological Science / William T. Keeton, James L. Gould
Yazar: Taner Tunç