
Memeli hormonlarının sayısı ve bunların işlevlerinin çeşitliliği çok büyüktür. Bu çeşitliliğe karşın, hücre içi iletişimde sadece çok sınırlı sayıda mekanizma rol oynar. Bu, somatik kontrol sağlayan bilgi iletişim stratejilerinden bazıları çok basittir, bazıları hücre zarıyla ilgili tartışmalarımızdan bilinmektedir ve bazıları da sadece hormonal kontrole özgüdür.
Hormonlar, hedef hücrelere çeşitli yollarla girerler. Steroyit hormonlarda olduğu gibi, bazıları hücre zarını ya doğrudan ya da bir reseptöre bağlanarak katederler. Bu reseptör kompleksi daha sonra, hormonun etkisini göstermesi için DNA’ya bağlanır. Az sayıda hormon hücre içine özel kanallar yoluyla girer. Bazıları da aktif olarak hücre içine taşınırlar.
Son olarak da birçok hormon hücreye hiç girmeyip bunun yerine hücre dışı reseptörlere bağlanırlar. Hücre dışı bağlanma, bir hücre içi etki yaratır. Bu, ya zarda bir iyon kanalının açılmasıyla ya da hücre içinde bir enzimi ya da ikinci haberciyi aktive ederek olur. Bu çeşitli mekanizmaların hücre metabolizmasını kontrol etmek üzere nasıl çalıştıklarını göreceğiz.
Hücre içinde de hormonal kontrol yöntemleri değişir. Testosteron gibi bazı hormonlar, DNA’ya bağlanırlar ve gen ifadesini doğrudan etkilerler ve sonuçta özgül enzimlerin yapımına yol açarlar. Adrenalin gibi diğer bazı hormonlar, mevcut enzimlerin aktivitesini kontrol ederler ya da etkilerini yapısal proteinleri değiştirerek gösterirler. Bir başka yöntem, farklı hormonların etkileşimini içerir; glukoz taşınması ve metabolizması üzerindeki zıt etkileriyle insulin ve glukagonda olduğu gibi.
Gen İfadesinin Düzenlenmesi
Steroyit hormonlar, örneğin böceklerdeki ekdizon ve omurgalılarda 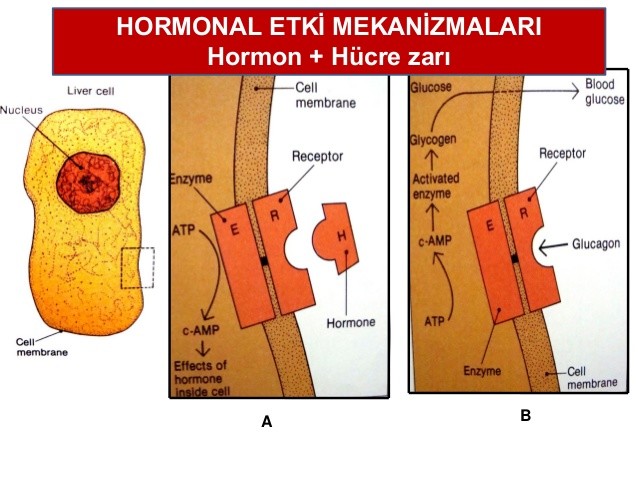 adrenal korteks ve gonatlar tarafından üretilen hormonlar, zarları kolaylıkla geçip hedef hücrenin sitoplazmasına girebilirler. Burada, steroyit (S) özgül bir stoplazmik reseptör proteine (R) bağlanır. Daha sonra kompleks, (S-R) çekirdeğe girer ve burada DNA’dan hangi RNA mesajlarının tercüme edilip (transkripsiyon) çekirdekten sitoplazmaya taşınacağını belirlemek yoluyla özgül genlerin aktivitelerinin düzenlenmesine yardımcı olur. Diğer bir ifadeyle hedef hücrenin genetik materyaliyle etkileşmek yoluyla steroyit hormonlar, çekirdekten ribozomlara protein sentezi (özellikle enzim sentezi) için hangi komutların gönderileceğini belirlerler. Zarlardan kolaylıkla geçip hücreye girebilen ve çekirdekte genlerin aktivitesini değiştiren ya da mitokondride enzim aktivitesini etkileyen ya da her ikisini birden yapan bir diğer hormon da tiroksindir.
adrenal korteks ve gonatlar tarafından üretilen hormonlar, zarları kolaylıkla geçip hedef hücrenin sitoplazmasına girebilirler. Burada, steroyit (S) özgül bir stoplazmik reseptör proteine (R) bağlanır. Daha sonra kompleks, (S-R) çekirdeğe girer ve burada DNA’dan hangi RNA mesajlarının tercüme edilip (transkripsiyon) çekirdekten sitoplazmaya taşınacağını belirlemek yoluyla özgül genlerin aktivitelerinin düzenlenmesine yardımcı olur. Diğer bir ifadeyle hedef hücrenin genetik materyaliyle etkileşmek yoluyla steroyit hormonlar, çekirdekten ribozomlara protein sentezi (özellikle enzim sentezi) için hangi komutların gönderileceğini belirlerler. Zarlardan kolaylıkla geçip hücreye girebilen ve çekirdekte genlerin aktivitesini değiştiren ya da mitokondride enzim aktivitesini etkileyen ya da her ikisini birden yapan bir diğer hormon da tiroksindir.
İkinci Haberciler
Siklik AMP’nin rolü konusunda 1960’larda hormonların hedef hücrelerini nasıl etkilediklerini anlama yönündeki girişimlerde heyecan verici ilerlemeler görüldü. Bu konuda önemli bir katkı, Western Reserve Üniversitesi’nden E.W. Sutherland ve T.W. Rall’den geldi. Bu araştırıcılar, glukagon ve adrenalinin karaciğer hücrelerini kana daha fazla glukoz salgılamak üzere uyarma mekanizmasını araştırıyorlardı. Bu hormonların, hücre içinde siklik adenozin monofosfat -kısaca siklik AMP ya da cAMP- adı verilen bir bileşiğin derişimini arttırdığını keşfettiler. Bu bileşiğin de glikojenin glukoza yıkımı için gerekli bir enzimin aktivasyonunu sağladığını buldular. Sonraki araştırmalar, glukagon ve adrenaline ek olarak, çok sayıda diğer hormonların, hedef hücrelerini cAMP derişimini arttırarak ya da azaltarak etkilediklerini ortaya koymuştur. Bu çalışma, şimdi Vanderbilt Üniversitesi’nde olan Sutherland’a 1971’de Nobel Ödülü kazandırmıştır.
Kuşkusuz, isminden tahmin ettiğiniz gibi, cAMP, ATP’yle (adenozin trifosfat) ilgili bir bileşiktir. Doğada çok yaygın olarak dağılmış olan cAMP, çalışılan hemen tüm hayvan dokularında (omurgalı ve omurgasız) ve bakterilerde 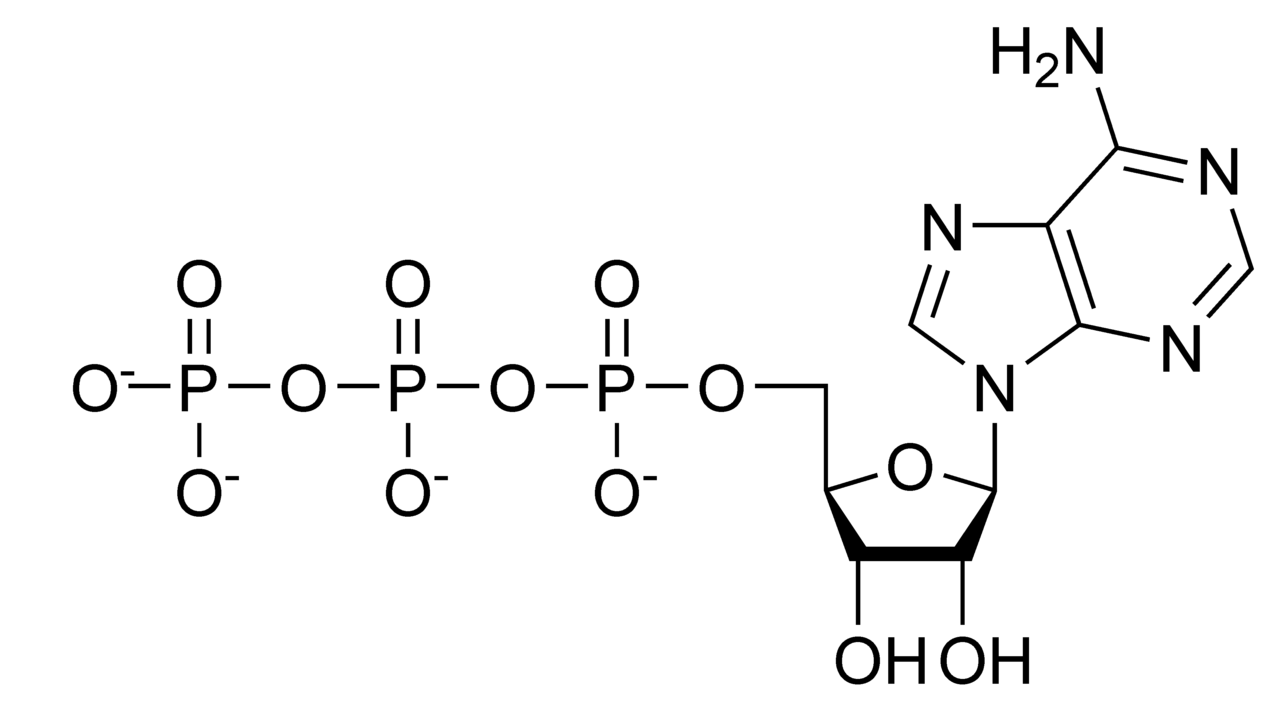 bulunmuştur. Canlı hücrede ATP’den, adenilat siklaz adı verilen ve hücre zarının içinde yerleşmiş olan bir enzimin katalizlediği bir reaksiyonla sentezlenir. Aralarında glukagon ve adrenalinin de bulunduğu birçok hormonun, aslında hedef hücrelere girmeyip hücre zarı üzerindeki reseptör bölgeleriyle zayıf bağlar yaptıkları yönünde kanıtlar artınca hormonal kontrolün ikinci haberci modeli önerildi. Bu modele göre, bir hücre dışı birinci haberci, hormonun kendisi, bir endokrin bezden hedef hücreye gider ve burada bir hücre içi ikinci habercinin, genellikle cAMP, yapımını uyarır. Biraz daha ayrıntılı olarak söylemek gerekirse; hormonun hedef hücrenin zarının dış yüzeyindeki oldukça özgül reseptör bölgesine bağlanması, adenilat siklazı aktive eder; bu da zarın iç yüzeyinde daha fazla cAMP yapımını katalizler. Daha sonra da miktarı artmış olan cAMP, sitoplazmik enzim sistemleriyle etkileşime girer ve hormonal uyarıma karşı hücrenin tipik tepkilerini başlatır. Diğer bir ifadeyle ilk hücre dışı sinyal (hormon ya da birinci haberci), hücrenin kimyasal düzeneğinin daha kolay anlayabileceği bir hücre içi sinyale (cAMP ya da ikinci haberci) dönüştürülür. Şimdi artık biliyoruz ki bir sinyal iletim proteini, üç birimli bir guanin bağlayıcı protein (G proteini), bu zincirde yer alır ve farklı hormon reseptörlerinin farklı etkiler göstermesini sağlar, örneğin siklazı inhibe etmek ya da farklı bir ikinci haberci sistemini aktive etmek gibi.
bulunmuştur. Canlı hücrede ATP’den, adenilat siklaz adı verilen ve hücre zarının içinde yerleşmiş olan bir enzimin katalizlediği bir reaksiyonla sentezlenir. Aralarında glukagon ve adrenalinin de bulunduğu birçok hormonun, aslında hedef hücrelere girmeyip hücre zarı üzerindeki reseptör bölgeleriyle zayıf bağlar yaptıkları yönünde kanıtlar artınca hormonal kontrolün ikinci haberci modeli önerildi. Bu modele göre, bir hücre dışı birinci haberci, hormonun kendisi, bir endokrin bezden hedef hücreye gider ve burada bir hücre içi ikinci habercinin, genellikle cAMP, yapımını uyarır. Biraz daha ayrıntılı olarak söylemek gerekirse; hormonun hedef hücrenin zarının dış yüzeyindeki oldukça özgül reseptör bölgesine bağlanması, adenilat siklazı aktive eder; bu da zarın iç yüzeyinde daha fazla cAMP yapımını katalizler. Daha sonra da miktarı artmış olan cAMP, sitoplazmik enzim sistemleriyle etkileşime girer ve hormonal uyarıma karşı hücrenin tipik tepkilerini başlatır. Diğer bir ifadeyle ilk hücre dışı sinyal (hormon ya da birinci haberci), hücrenin kimyasal düzeneğinin daha kolay anlayabileceği bir hücre içi sinyale (cAMP ya da ikinci haberci) dönüştürülür. Şimdi artık biliyoruz ki bir sinyal iletim proteini, üç birimli bir guanin bağlayıcı protein (G proteini), bu zincirde yer alır ve farklı hormon reseptörlerinin farklı etkiler göstermesini sağlar, örneğin siklazı inhibe etmek ya da farklı bir ikinci haberci sistemini aktive etmek gibi.
Karaciğerde glukagonun ya da adrenalinin etkisiyle glukoz üretiminin artışı, artan cAMP miktarının hücreyi etkilemesine bir örnektir. Burada cAMP, protein kinaz adı verilen bir enzim grubunu aktive eder. Aktive olan protein kinaz da ikinci bir enzimi, bu da üçüncü bir enzirni aktive eder. Bu enzim ise glikojenin glukoza yakınındaki ilk reaksiyonu katalizler. Bu, enzimce kataliz edilen reaksiyonlar zinciri, kademeli dizi reaksiyonlara bir  örnek olup başlangıçtaki hormon bağlarna olayının etkilerinin büyük ölçüde amplifikasyonunu sağlar. Bir enzim, tekrar tekrar kullanılabildiği için bir tek aktif adenilat siklaz molekülü, yaklaşık 100 cAMP molekülünün yapımını katalizleyebilir. Yapılan bir cAMP molekülü de kabaca 100 enzim A molekülünün yapımını başlatabilir, vb. Bunun net sonucu olarak bir tek glukagon ya da adrenalin molekülü, sadece bir ya da iki dakika içinde 1010 (10 milyar) glukoz molekülünün oluşmasını sağlayabilir. Böylece iletişimin neden çok az hormon miktarı gerektirdiğini de görmüş oluyoruz. Yukarıdaki reaksiyonlar dizisi bir hedef hücrenin glukagon ya da adrenalin tarafından uyarılmasının tek sonucu değildir. Bu hormonlarla ilgili önceki tartışmalarımızdan hatırlayacağınız gibi, glukoz serbestleşmesi, bu hormonların kan şekeri düzeyini arttırmalarını sağlayan çeşitli yolların sadece bir tanesidir. Bunların önemli etkilerinden bir başkası, glikojen sentezini inhibe etmeleridir. Bu etki, şimdi tartıştığımız etki gibi, cAMP tarafı ndan aktive edilen protein kinaza bağımlıdır. Daha önce glukoz yapımı için gerekli bir enzimi aktive ettiğini gördüğümüz protein kinaz (enzim A), burada bir enzimi inaktive etmekte ve böylece glukozun glikojene dönüşmesini engellemektedir. Protein kinazın adenilat siklaz sisteminin aracılık yaptığı tüm hormonal etkilerde temel bağlayıcı faktör olduğu yolunda tatmin edici miktarda kanıt vardır. Hem glukagonun hem de adrenalinin incelediğimiz olayı başlatabildiklerini gördük. Bu iki hormon, aslında kimyasal olarak oldukça farklıdır. Adenilat siklaz üzerinde nasıl oluyor da aynı etkiyi yapıyorlar? Cevap şudur: Her ne kadar ikisi de zar üzerinde farklı reseptör moieküllere bağlanıyorlarsa da her iki tip reseptör de adenilat siklazı aktive etme kapasitesine sahiptir. Böylece iki hormon aynı etkiyi gösterirler. Fakat bu, sadece karaciğer için doğrudur; bu iki horrnonun kas üzerindeki etkisi aynı değildir. Adenilat siklaz yoluyla etki eden adrenalin, kas hücrelerinin glikojenden glukoz yapımını uyarabilir; fakat glukagon bunu yapamaz çünkü kas hücrelerinin zarında glukagon reseptörleri yoktur.
örnek olup başlangıçtaki hormon bağlarna olayının etkilerinin büyük ölçüde amplifikasyonunu sağlar. Bir enzim, tekrar tekrar kullanılabildiği için bir tek aktif adenilat siklaz molekülü, yaklaşık 100 cAMP molekülünün yapımını katalizleyebilir. Yapılan bir cAMP molekülü de kabaca 100 enzim A molekülünün yapımını başlatabilir, vb. Bunun net sonucu olarak bir tek glukagon ya da adrenalin molekülü, sadece bir ya da iki dakika içinde 1010 (10 milyar) glukoz molekülünün oluşmasını sağlayabilir. Böylece iletişimin neden çok az hormon miktarı gerektirdiğini de görmüş oluyoruz. Yukarıdaki reaksiyonlar dizisi bir hedef hücrenin glukagon ya da adrenalin tarafından uyarılmasının tek sonucu değildir. Bu hormonlarla ilgili önceki tartışmalarımızdan hatırlayacağınız gibi, glukoz serbestleşmesi, bu hormonların kan şekeri düzeyini arttırmalarını sağlayan çeşitli yolların sadece bir tanesidir. Bunların önemli etkilerinden bir başkası, glikojen sentezini inhibe etmeleridir. Bu etki, şimdi tartıştığımız etki gibi, cAMP tarafı ndan aktive edilen protein kinaza bağımlıdır. Daha önce glukoz yapımı için gerekli bir enzimi aktive ettiğini gördüğümüz protein kinaz (enzim A), burada bir enzimi inaktive etmekte ve böylece glukozun glikojene dönüşmesini engellemektedir. Protein kinazın adenilat siklaz sisteminin aracılık yaptığı tüm hormonal etkilerde temel bağlayıcı faktör olduğu yolunda tatmin edici miktarda kanıt vardır. Hem glukagonun hem de adrenalinin incelediğimiz olayı başlatabildiklerini gördük. Bu iki hormon, aslında kimyasal olarak oldukça farklıdır. Adenilat siklaz üzerinde nasıl oluyor da aynı etkiyi yapıyorlar? Cevap şudur: Her ne kadar ikisi de zar üzerinde farklı reseptör moieküllere bağlanıyorlarsa da her iki tip reseptör de adenilat siklazı aktive etme kapasitesine sahiptir. Böylece iki hormon aynı etkiyi gösterirler. Fakat bu, sadece karaciğer için doğrudur; bu iki horrnonun kas üzerindeki etkisi aynı değildir. Adenilat siklaz yoluyla etki eden adrenalin, kas hücrelerinin glikojenden glukoz yapımını uyarabilir; fakat glukagon bunu yapamaz çünkü kas hücrelerinin zarında glukagon reseptörleri yoktur.
Bu glukoz düzenlenmesi örneğinden çeşitli önemli sonuçlar çıkartabiliriz. Birincisi, belirli bir hücre tipinin bir hormondan etkilenmesi -yani o  hormonun hedefi olup olmadığı- hücre zarında reseptörlerin bulunup bulunmamasına bağlıdır. İkincisi, eğer bir hücrede farklı hormonlar için siklaz aktive edici reseptörler varsa o hücre, iki ya da daha fazla farklı hormona aynı tepkiyi gösterir. Hormonlar sadece reseptörler için özgüldürler; bağlanmalarının, gproteini ve ikinci haberci aktivasyonu aracılığıyla başlattığı reaksiyonlar için değil. Hormon reseptörlerinin hücreye özgül dağılımı, adenilat siklaz sistemine bağımlı birçok farklı hormonun (glukagon ve adrenaline ek olarak gastrin, sekretin, paratiroyit hormon, kalsitonin, ön hipofizin tropik hormonları ve hipotalamustan salgılanan salgılatıcı hormonların en azından bazıları), her hücredeki ilk etkilerinin adenilat siklazın aktivasyonu olmasına karşın, nasıl kendilerine özgü hedef hücreleri olduğunu açıklamaya yardımcı olur. Cevaplaması çok daha zor olan soru ise farklı hücrelerin cAMP içeriklerindeki değişikliklere neden bu kadar farklı tepkiler gösterdikleridir. cAMP artışının neden tiroyit hücrelerinde tiroksin yapımında, adrenal korteks hücrelerinde kortizon yapımında, karaciğer hücrelerinde glukoz yapımında ve uterus hücrelerinde protein sentezinde artışa yol açması gerektiği hiçbir biçimde açıklanamaz. İki varsayım yapılabilir. Birincisi, hücrede çok sayıda farklı kimyasal olayın cAMP tarafından düzenleniyor olmasıdır ki bu varsayırm destekleyen bol miktarda bulgu vardır. İkinci varsayım, bir hücrenin değişen cAMP derişimine vereceği tepki, onun kendi kimyasal bileşimine bağlıdır. Tiroyit, adrenal, karaciğer ve uterus hücreleri farklı kimyasal profillere sahiptirler. Böylece hormonun yol açtığı cAMP artışı, bu dört hücre tipinin her birinde farklı enzim sistemlerini etkileyecek ve artışın etkileri de buna göre farklı olacaktır.
hormonun hedefi olup olmadığı- hücre zarında reseptörlerin bulunup bulunmamasına bağlıdır. İkincisi, eğer bir hücrede farklı hormonlar için siklaz aktive edici reseptörler varsa o hücre, iki ya da daha fazla farklı hormona aynı tepkiyi gösterir. Hormonlar sadece reseptörler için özgüldürler; bağlanmalarının, gproteini ve ikinci haberci aktivasyonu aracılığıyla başlattığı reaksiyonlar için değil. Hormon reseptörlerinin hücreye özgül dağılımı, adenilat siklaz sistemine bağımlı birçok farklı hormonun (glukagon ve adrenaline ek olarak gastrin, sekretin, paratiroyit hormon, kalsitonin, ön hipofizin tropik hormonları ve hipotalamustan salgılanan salgılatıcı hormonların en azından bazıları), her hücredeki ilk etkilerinin adenilat siklazın aktivasyonu olmasına karşın, nasıl kendilerine özgü hedef hücreleri olduğunu açıklamaya yardımcı olur. Cevaplaması çok daha zor olan soru ise farklı hücrelerin cAMP içeriklerindeki değişikliklere neden bu kadar farklı tepkiler gösterdikleridir. cAMP artışının neden tiroyit hücrelerinde tiroksin yapımında, adrenal korteks hücrelerinde kortizon yapımında, karaciğer hücrelerinde glukoz yapımında ve uterus hücrelerinde protein sentezinde artışa yol açması gerektiği hiçbir biçimde açıklanamaz. İki varsayım yapılabilir. Birincisi, hücrede çok sayıda farklı kimyasal olayın cAMP tarafından düzenleniyor olmasıdır ki bu varsayırm destekleyen bol miktarda bulgu vardır. İkinci varsayım, bir hücrenin değişen cAMP derişimine vereceği tepki, onun kendi kimyasal bileşimine bağlıdır. Tiroyit, adrenal, karaciğer ve uterus hücreleri farklı kimyasal profillere sahiptirler. Böylece hormonun yol açtığı cAMP artışı, bu dört hücre tipinin her birinde farklı enzim sistemlerini etkileyecek ve artışın etkileri de buna göre farklı olacaktır.
Hormonlar dışında çeşitli kimyasallar da hücreleri, cAMP düzeyini değiştirerek etkilerler. Bunlardan bazıları, örneğin histamin, adenilat siklaz yoluyla çalışırken diğerleri, cAMP’yi parçalayan enzim olan fosfodiesteraz üzerinde etkili olurlar. Normal bir hücrede, cAMP derişimini kontrol altında tutan bu enzimden her zaman belli bir miktar bulunur. Böylece, eğer adrenalin cAMP derişiminde bir artışa yol açarsa, hormon ortadan kalkar kalkmaz fosfodiesteraz normal düzeye geri dönüşü sağlar. Bu enzimi inhibe edebilen bir kimyasalın, örneğin kafein, ya da bu enzimi aktive edebilen bir kimyasalın, örneğin nikotin, hücrede cAMP düzeylerini önemli ölçüde etkileyeceği gayet açıktır.
İlk keşfedildiğinde cAMP’nin ikinci haberci olarak oynadığı rol bazı araştırıcıları o kadar etkilemiştir ki bunlar, cAMP’nin hormonların ve diğer hormon benzeri bileşiklerin etkilerinde muhtemel bir evrensel aracı olduğunu düşünmüşlerdir. Kısa bir süre sonra bu fikrin doğru olmadığı gösterildi. Örneğin insulinin (etkisi glukagonunkine ve adrenalininkine antagonisttir) temel etkisi cAMP derişiminde bir değişiklik değil; fakat doğrudan hücre içi enzimleri etkilemektir.
Çeşitli başka kontrol kimyasalları (özellikle sinir sisteminde asetilkolin ve ANF hormonu) GTP’yi cGMP’ye çevirerek etki ederler. Fakat cGMP ikinci haberci sisteminin ayrıntıları henüz tam anlaşılmış değildir.
İkinci haberci olarak iyonlar, daha önce söylediğimiz gibi, hücre metabolizması üzerindeki etkilerini hücrenin içine girmeden gösteren hormonlar, bunu ya bir ikinci haberciyi aktive ederek, ya da bir iyon 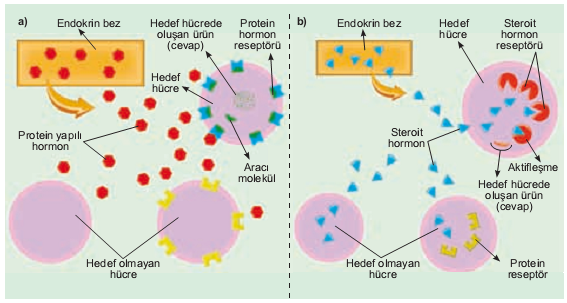 kanalını açarak başarırlar. Gerçekte, cAMP sistemi ve iyon kanalı stratejisi o kadar yakından bağlıdır ki bu iyonları da ikinci haberciler olarak düşünmek mantıklı olacaktır. Şimdiye kadar en yaygın olarak bilinen ikinci haberci iyon Ca++’dur. Kalsiyum iyonlarının bu işlevi görmesini sağlayan, Ca++’un sitosoldeki derişiminin normalde düşük tutulmasıdır: Ca++ iyonları hem ER içine, hem de hücre dışına aktif olarak pompalandıkları gibi mitokondri tarafından alınıp depolanırlar ve sitosolde serbest olarak bulunan çeşitli moleküllere bağlanırlar. Bunun sonucunda bir hormon, özgül reseptörüne bağlanıp zarda bir Ca++ kanalını açınca güçlü elektrokimyasal gradiyent nedeniyle Ca++ iyonları içeri hücum ederler. Diğer hormonlar, bir lipit ikinci haberciyi –inozitol trifosfat- aktive ederek (G-proteini kompleksi yoluyla) etki ederler. Bu ikinci haberci, ER’ye difüze olarak buradaki kapılı Ca++ kanallarının açılmasını sağlar. Her iki halde de Ca++ iyonları bazı hücre içi enzimlere bağlanıp bunları aktive ederler. Bunlardan en yaygın olarak bulunan ve en iyi bilineni, 148 amino asitten oluşan kalmodulindir. Ca++- kalmodulin kompleksi, daha sonra kontrol edilmekte olan sistemin enzimlerini aktive eder. Bu hücrede hangi enzimlerin düzenleneceği, o hücrenin tipine, diğer bir ifadeyle, o hücrede aktif olan genlere bağlıdır. cAMP gibi kalmodulin de farklı hücrelerde farklı enzim gruplarını kontrol eder. İyon kanalı ve Ca++- kalmodulin kompleksinin işleyişi kas hücrelerinde görülebilir. Adrenalin, kas hücrelerini aktiviteye hazırlamak için glikojenin glukoza yıkımını uyarırken sadece cAMP’yi değil Ca++’u da ikinci haberci olarak kullanır. Adrenalinin özgül reseptörüne bağlanması, bir Ca++ kanalını açar, sonuçta oluşan kalmodulin kompleksi fosforilaz kinaza bağlanır. Bu enzim ise, glikojeni glukoz -1- fosfata dönüştürüp bunu glikolitik yola gönderen enzim olan glikojen fosforilazı aktive eder. Böylece kasta, glukoz yapımını uyarmak için hem cAMP hem de aktive edilmiş kalmodulin gereklidir; aktive edilmiş kalmodulinin fosforilaz kinaza bağlanması, kinazın aktivasyonunun bir kısmıdır. cAMP, aktif kalmodulin ve çeşitli enzimler arasındaki bu etkileşim, karmaşık organizmaların kimyasını oluşturan bağlantılar hakkında bize bir ipucu verir.
kanalını açarak başarırlar. Gerçekte, cAMP sistemi ve iyon kanalı stratejisi o kadar yakından bağlıdır ki bu iyonları da ikinci haberciler olarak düşünmek mantıklı olacaktır. Şimdiye kadar en yaygın olarak bilinen ikinci haberci iyon Ca++’dur. Kalsiyum iyonlarının bu işlevi görmesini sağlayan, Ca++’un sitosoldeki derişiminin normalde düşük tutulmasıdır: Ca++ iyonları hem ER içine, hem de hücre dışına aktif olarak pompalandıkları gibi mitokondri tarafından alınıp depolanırlar ve sitosolde serbest olarak bulunan çeşitli moleküllere bağlanırlar. Bunun sonucunda bir hormon, özgül reseptörüne bağlanıp zarda bir Ca++ kanalını açınca güçlü elektrokimyasal gradiyent nedeniyle Ca++ iyonları içeri hücum ederler. Diğer hormonlar, bir lipit ikinci haberciyi –inozitol trifosfat- aktive ederek (G-proteini kompleksi yoluyla) etki ederler. Bu ikinci haberci, ER’ye difüze olarak buradaki kapılı Ca++ kanallarının açılmasını sağlar. Her iki halde de Ca++ iyonları bazı hücre içi enzimlere bağlanıp bunları aktive ederler. Bunlardan en yaygın olarak bulunan ve en iyi bilineni, 148 amino asitten oluşan kalmodulindir. Ca++- kalmodulin kompleksi, daha sonra kontrol edilmekte olan sistemin enzimlerini aktive eder. Bu hücrede hangi enzimlerin düzenleneceği, o hücrenin tipine, diğer bir ifadeyle, o hücrede aktif olan genlere bağlıdır. cAMP gibi kalmodulin de farklı hücrelerde farklı enzim gruplarını kontrol eder. İyon kanalı ve Ca++- kalmodulin kompleksinin işleyişi kas hücrelerinde görülebilir. Adrenalin, kas hücrelerini aktiviteye hazırlamak için glikojenin glukoza yıkımını uyarırken sadece cAMP’yi değil Ca++’u da ikinci haberci olarak kullanır. Adrenalinin özgül reseptörüne bağlanması, bir Ca++ kanalını açar, sonuçta oluşan kalmodulin kompleksi fosforilaz kinaza bağlanır. Bu enzim ise, glikojeni glukoz -1- fosfata dönüştürüp bunu glikolitik yola gönderen enzim olan glikojen fosforilazı aktive eder. Böylece kasta, glukoz yapımını uyarmak için hem cAMP hem de aktive edilmiş kalmodulin gereklidir; aktive edilmiş kalmodulinin fosforilaz kinaza bağlanması, kinazın aktivasyonunun bir kısmıdır. cAMP, aktif kalmodulin ve çeşitli enzimler arasındaki bu etkileşim, karmaşık organizmaların kimyasını oluşturan bağlantılar hakkında bize bir ipucu verir.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç