
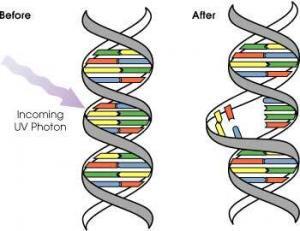
 DNA, nükleotid adı verilen yapı birimlerinden oluşur. Nükleotitler art arda dizilerek DNA zincirini oluşturur. DNA, birbiriyle nükleotitlerdeki bazlar aracılığıyla eşleşebilen iki zincirin eşleşip oluşturduğu sarmal bir yapıdır. Nükleotitlerin yapısında; azotlu bir baz, deoksiriboz şekeri ve fosfat grubu vardır. Azotlu baz; adenin (A), guanin (G), sitozin (C) ya da timin (T) olabilir. Adenin timin bazıyla, sitozin de normal koşullarda guanin bazıyla eşleşir. Bu eşleşmelerin dışında bir eşleşme olursa DNA’da hasar meydana gelmiş olur (mutasyon) ve hücrenin yaşamına devam edebilmesi için bu hasarı onarması gerekir. Bunun dışında, bazlara kimyasal grupların eklenmesi ya da bazların kopması da DNA hasarına yol açar.
DNA, nükleotid adı verilen yapı birimlerinden oluşur. Nükleotitler art arda dizilerek DNA zincirini oluşturur. DNA, birbiriyle nükleotitlerdeki bazlar aracılığıyla eşleşebilen iki zincirin eşleşip oluşturduğu sarmal bir yapıdır. Nükleotitlerin yapısında; azotlu bir baz, deoksiriboz şekeri ve fosfat grubu vardır. Azotlu baz; adenin (A), guanin (G), sitozin (C) ya da timin (T) olabilir. Adenin timin bazıyla, sitozin de normal koşullarda guanin bazıyla eşleşir. Bu eşleşmelerin dışında bir eşleşme olursa DNA’da hasar meydana gelmiş olur (mutasyon) ve hücrenin yaşamına devam edebilmesi için bu hasarı onarması gerekir. Bunun dışında, bazlara kimyasal grupların eklenmesi ya da bazların kopması da DNA hasarına yol açar.
Organizmalar, yaşamları boyunca DNA’larını hasara uğratan içsel ve çevresel faktörlere maruz kalırlar. Eğer DNA’daki hasar onarılamazsa; mutasyonla, hastalıkla ve hücre ölümüyle sonuçlanabilir. DNA hasarına hücresel cevap, DNA hasar kontrol yolağının aktivasyonu ya da DNA hasarının doğrudan onarımıdır. Baz modifikasyonları, DNA zincir kırıkları, zincirlerin çapraz bağlanması ve yanlış eşleşmeler gibi çeşitli DNA hasarı türleri mevcuttur. Ayrıca çok sayıda DNA onarım yolağı vardır. Her bir onarım yolağı belli bir hasar türüyle bağlantılıdır. Bir tür hasar birden fazla onarım mekanizması tarafından da hedeflenebilir. En önemli DNA onarım mekanizmaları; hatalı eşleşmelerin onarımı (MMR), nükleotid çıkarma onarımı (NER), baz çıkarma onarımı (BER), homolog rekombinasyon onarımı (HR) ve homolog olmayan onarımdır (NHEJ). Bu yolaklar çok sayıda proteinin varlığını gerektirir. Bunun aksine, alkil grubu eklenmiş bir guanin bazı, sadece bir metil transferaz enzimi olan MGMT tarafından onarılabilir. MGMT bazdaki alkil grubunu kendi yapısındaki sistein (enzimin yapısında bulunan bir amino asit) kalıntılarına transfer eder. Fotoliyaz enzimi de UV radyasyon tarafından üretilen primidin dimerleri (yan yana iki timin ya da sitozin nükleotidi arasında bağ oluşması) arasındaki kovalent bağları ayırabilir. Bu enzim UV’nin hasar verdiği yere bağlanır ve aktivite göstermek için 350-450 nm dalgaboyunda ışığa enerji kaynağı olarak gereksinim duyar. Başka bir NER bağımsız yolak, UV’nin yol açtığı hasarı onarabilir. Bu yolağa UVER denir Saccaromyces pombe gibi mayalar dahil birkaç organizmada görülür. UVER’deki anahtar faktör DNA’da hasarlı bölgeyi kesen endonükleaz Uve1/UVDE’dir.
 Ülkemizde doğmuş ve günümüzde çalışmalarını Amerika Birleşik Devletleri’nde sürdüren bilim insanı Aziz Sancar, Nobel ödülü kazandığı çalışmada, UV’nin DNA’da yol açtığı hasarın onarım mekanizmasını göstermiştir. Bu çalışmada Aziz Sancar, bakterileri ölümcül dozda UV radyasyona maruz bıraktıktan sonra, görünür mavi ışık altında bir süre sonra iyileştiklerini gözlemdi. 1976 yılında doktora tezinde, UV kaynaklı DNA hasarını onaran fotoliyaz adlı enzimi saflaştırmayı başardı. Bakterilerde ışığa bağlı onarım sisteminin dışında, karanlıkta da işlev gören bir mekanizma keşfedildi. Önceki çalışması fotoliyazda olduğu gibi Sancar, karanlıkta olan sistemin moleküler mekanizmasını araştırdı. Birkaç yıl içerisinde, uvrA, uvrB ve uvrC genlerinin kodladığı enzimleri tanımladı ve izole etti. Yaptığı deneylerde bu enzimlerin DNA hasarlarını tanımlayabildiğini, DNA’da iki tane nükleotid çıkarma işlemi yaptığını ve bu işlemlerden birinin de hasarlı bölgede olduğunu gösterdi. Fotoliyaz konusunda ise, fotoliyazın biyolojik saati düzenlemeye yardımcı olduğunu gösteren çalışmaya dahil oldu.
Ülkemizde doğmuş ve günümüzde çalışmalarını Amerika Birleşik Devletleri’nde sürdüren bilim insanı Aziz Sancar, Nobel ödülü kazandığı çalışmada, UV’nin DNA’da yol açtığı hasarın onarım mekanizmasını göstermiştir. Bu çalışmada Aziz Sancar, bakterileri ölümcül dozda UV radyasyona maruz bıraktıktan sonra, görünür mavi ışık altında bir süre sonra iyileştiklerini gözlemdi. 1976 yılında doktora tezinde, UV kaynaklı DNA hasarını onaran fotoliyaz adlı enzimi saflaştırmayı başardı. Bakterilerde ışığa bağlı onarım sisteminin dışında, karanlıkta da işlev gören bir mekanizma keşfedildi. Önceki çalışması fotoliyazda olduğu gibi Sancar, karanlıkta olan sistemin moleküler mekanizmasını araştırdı. Birkaç yıl içerisinde, uvrA, uvrB ve uvrC genlerinin kodladığı enzimleri tanımladı ve izole etti. Yaptığı deneylerde bu enzimlerin DNA hasarlarını tanımlayabildiğini, DNA’da iki tane nükleotid çıkarma işlemi yaptığını ve bu işlemlerden birinin de hasarlı bölgede olduğunu gösterdi. Fotoliyaz konusunda ise, fotoliyazın biyolojik saati düzenlemeye yardımcı olduğunu gösteren çalışmaya dahil oldu.
Hatalı Eşleşme Onarımı
MMR’nin ana görevi, yanlış baz eşleşmelerini ve replikasyon sırasında meydana gelen küçük parça eklenmelerini ya da eksilmelerini düzenlemektir. İnsan hücrelerinde MMR mekanizmasında, yanlış baz eşleşmeleri, MSH2-MSH6 adlı protein kompleksi tarafından tanınır. Küçük nükleotid eklemeleri ve eksilmeleri MSH2-MSH6 ve MSH2-MSH3 protein kompleksleri tarafından tanımlanır. Ökaryot hücrelerinde (bitkilerde ve memelilerde bulunan hücre tipi) eski ve yeni sentezlenen DNA zinciri arasındaki farkın nasıl ayırt edildiği henüz anlaşılamamıştır. İplik ayrımı PCNA isimli bir faktör tarafından yapılabilir. Ya da yeni sentezlenen zincirin boşlukları, kesikleri ve serbest ucu tanınıyor olabilir. Daha sonraki aşamada yeni sentezlenen zincir parçalanarak, yanlış eşleşme yok edilir.
Nükleotid Çıkarma Onarımı
 UV hasarları dahil çok çeşitli DNA hasarlarını NER yok edebilir. NER iki tane alt yolak içerir. Bunlar: global genom onarımı (GGR) ve transkripsiyonla eşleşen onarım (TCR)’dir. TCR, özgül olarak aktif genin transkribe ona zincirini (RNA sentezinin yapıldığı zincir) yeniler. GGR ve TCR arasındaki temel fark, hasarı tanıyan proteinlerin farklı olmasıdır. DDB1, DDB2 ve XPC-hHR23B proteinleri GGR’nin tanıma sistemi içindedir. TCR mekanizması ise, RNA polimeraz II tarafından başlatılır. Ayrıca TCR için, CSA ve CSB faktörleri de gereklidir. GGR ve TCR’de daha sonraki aşamalarda yer alan proteinler benzerdir. IIH adlı 9 alt birimden oluşan bir transkripsiyon faktörü DNA’da hasarlı bölgeyi tanır. Bu aşamada başlangıç tanıma faktörleri DNA’dan ayrılır. IIH’nin iki alt birimi, XBP ve XPD helikaz aktivitesi gösterirler ve DNA zincirini hasar etrafında çözerler.
UV hasarları dahil çok çeşitli DNA hasarlarını NER yok edebilir. NER iki tane alt yolak içerir. Bunlar: global genom onarımı (GGR) ve transkripsiyonla eşleşen onarım (TCR)’dir. TCR, özgül olarak aktif genin transkribe ona zincirini (RNA sentezinin yapıldığı zincir) yeniler. GGR ve TCR arasındaki temel fark, hasarı tanıyan proteinlerin farklı olmasıdır. DDB1, DDB2 ve XPC-hHR23B proteinleri GGR’nin tanıma sistemi içindedir. TCR mekanizması ise, RNA polimeraz II tarafından başlatılır. Ayrıca TCR için, CSA ve CSB faktörleri de gereklidir. GGR ve TCR’de daha sonraki aşamalarda yer alan proteinler benzerdir. IIH adlı 9 alt birimden oluşan bir transkripsiyon faktörü DNA’da hasarlı bölgeyi tanır. Bu aşamada başlangıç tanıma faktörleri DNA’dan ayrılır. IIH’nin iki alt birimi, XBP ve XPD helikaz aktivitesi gösterirler ve DNA zincirini hasar etrafında çözerler.
Hasarlı bölgeye bağlanan diğer faktörler XPG ve XPA-RPA’dır. XPA-RPA, NER kompleksinin doğru bir şekilde kurulup kurulmadığını kontrol eder ve hasarlı DNA’nın doğru şekilde kesildiğinden emin olur. XPF-ERCC1’in bağlanmasının ardından XPG ve XPF-ERCC1 tarafından çift çıkarılma gerçekleşir. Bu aşamada hasar, 24-32 nükleotid uzunluğunda uzun bir oligonükleotit olarak salınır. Onarım DNA sentezi tarafından tamamlanır. NER mekanizmasındaki hatadan kaynaklı olan hastalık, Xeroderma pigmentosum’dur. TCR’deki fonksiyon bozukluğu ise; Cockayne sendromu (CS) ve trichothiodystrophy (TTD) hastalıklarına yol açar.
Baz Çıkarma Onarımı
BER temel olarak; bazın alkillenmesi, oksidasyonu ve deaminasyonu (amin grubunun çıkarılması) sonucu ortaya çıkan hasarları onarır. Hücre birkaç çeşit DNA glikosilaz enzimi içerir. Bu enzimlerin her biri farklı bir molekülü hedefler. Nükleotit içerisinde, bazın şeker ile yaptığı bağın, DNA glikosilaz tarafından parçalanmasından sonra; hasarlı baz DNA’dan ayrılır ve DNA’da apürünik ya da apirimidik bir bölge (AP bölgesi) ortaya çıkar. AP bölgesi kendiliğinden de bir hasar olarak ortaya çıkabilir. Çift fonksiyonlu glikozilaz enzimleri, AP bölgesindeki şeker-fosfat bağını kesen bir aktiviteye de sahiptir. Sonuç olarak parçalanmış şeker bakiyesi, AP endonükleaz ya da DNA polimeraz beta enzimi tarafından parçalanır. Bir nükleotitlik boşluk, DNA polimeraz beta tarafından doldurulur ve bağlanır. AP bölgelerinin işlenmesi, monofonksiyonel DNA glikozilaz tarafından yapılır ve bir AP nükleaz tarafından DNA’nın 5′ ucunun kesilmesini gerektirir. Polimeraz beta bölgeye bir nükleotit ekler ve onun deoksiribofosfodiesteraz aktivitesi 5’ucunda küçük bir parçayı yok eder. Kalan boşluk kapatılır. BER yolağında hasarlı nükleotide ek olarak 2-8 nükleotit ortadan kaldırılır. Günümüzde BER’deki fonksiyon bozukluğundan kaynaklı herhangi bir hastalık bilinmemektedir. Buna rağmen, DNA glikozilaz enzimlerinin aktivitesi eksik olan transgenik farede, embriyo ölüme gitmiştir. Ancak, enzimlerden sadece birinde mutasyon olması, farede herhangi bir anormaliye yol açmamıştır.
Homolog Rekombinasyon Onarımı
 DNA’daki çift iplik kırıkları, HR ya da NHEJ mekanizmalarıyla onarılabilir. HR, kalıp olarak homolog kromozomdaki (ökaryot hücrelerde her kromozomdan iki tane vardır) DNA kalıbını kullanır ve bu yüksek oranda doğruluk sağlar. Bunun aksine NHEJ, kalıp kullanmadan kırılmış uçları tekrar bir araya getirir ve bu mekanizma sıklıkla bazı nükleotitlerin kaybıyla sonuçlanır. Bu nedenle NHEJ mekanizması hataya eğilimlidir. Her bir yolağın göreceli katılımı, hücrenin hangi evrede olduğuna bağlıdır. NHEJ G1 fazında (hücre çoğalmasında mitoz bölünmenin bir aşaması) daha aktiftir ve HR ise S ve G2 fazlarında (mitoz aşamaları) aktiftir. HR onarımı sırasında çift iplik kırıkları, 3′ uçlu (DNA’da fosfatın serbest olduğu uç) tek zincirlere dönüştürülür ve bu zincirler RPA’ya bağlanır. Çift iplik kırığının işlenmesi, MRE11-RAD50-NBS1’i gerektirir. RAD52 NPA ile ilişki kurar ve RAD51’in tek zincirli DNA’ya bağlanmasını teşvik eder. DNA sentezinden sonra zincirler arasında değişim olur.
DNA’daki çift iplik kırıkları, HR ya da NHEJ mekanizmalarıyla onarılabilir. HR, kalıp olarak homolog kromozomdaki (ökaryot hücrelerde her kromozomdan iki tane vardır) DNA kalıbını kullanır ve bu yüksek oranda doğruluk sağlar. Bunun aksine NHEJ, kalıp kullanmadan kırılmış uçları tekrar bir araya getirir ve bu mekanizma sıklıkla bazı nükleotitlerin kaybıyla sonuçlanır. Bu nedenle NHEJ mekanizması hataya eğilimlidir. Her bir yolağın göreceli katılımı, hücrenin hangi evrede olduğuna bağlıdır. NHEJ G1 fazında (hücre çoğalmasında mitoz bölünmenin bir aşaması) daha aktiftir ve HR ise S ve G2 fazlarında (mitoz aşamaları) aktiftir. HR onarımı sırasında çift iplik kırıkları, 3′ uçlu (DNA’da fosfatın serbest olduğu uç) tek zincirlere dönüştürülür ve bu zincirler RPA’ya bağlanır. Çift iplik kırığının işlenmesi, MRE11-RAD50-NBS1’i gerektirir. RAD52 NPA ile ilişki kurar ve RAD51’in tek zincirli DNA’ya bağlanmasını teşvik eder. DNA sentezinden sonra zincirler arasında değişim olur.
Homolog Olmayan Onarım
NHEJ, Ku70-Ku80 protein kompleksinin DNA uçlarına bağlanmasıyla başlar. Daha yüksek ökaryotlarda DNA protein kinazın katalitik alt birimi, sonradan dahil olur. Bu aşamada DNA polimeraz gerekli olabilir. Son aşamada DNA uçları, XRCC4-DNA ligaz ile yeniden bağlanır.
Çift iplik kırıklarının hatalı onarımı; kromozomların yapısal kararsızlığıyla ve bu da kromozomların yeniden bir araya gelmesi ya da kromozom kaybıyla sonuçlanır. Ataxia talengiectasia (AT), Nijmegen kırılma sendromu (NBS), meme ve yumurtalık kanserine neden olan BRCA1 ve BRCA2 genlerindeki mutasyonlar çift iplik kırığı onarımındaki hatalarla ilgilidir. Fakat bu sendromlar; HR ya da NHEJ’in inaktivasyonundan çok, çift iplik kırılmasındaki hatalardan kaynaklanır.
DNA onarım mekanizmalarının daha iyi anlaşılması; mutasyonların nasıl meydana geldiğini ve kanser başta olmak üzere genetik hastalıkların tedavisi için DNA’yı nasıl manipüle edebileceğimizi anlamamızı sağlar.
Kaynakça:
https://www.nobelprize.org/nobel_prizes/chemistry/laureates/2015/popular-chemistryprize2015.pdf
Fleck and Nielsen. DNA repair. Journal of Cell Science 117, 515-517
Yazar: Ayça Olcay