
Yakın zamana kadar bilim adamlarının çoğu, karmaşık davranışların (çeşitli duyulan entegre eden davranışlar ve çeşitli kasların sırasıyla koordineli bir şekilde hareket ettirilmesiyle ortaya çıkan davranışlar) birbiriyle etkileşen refleks arkları aracılığıyla meydana geldiğini zannederlerdi. Her refleks arkının tek bir duyu aldığı ve tek bir kası (ya da eli geri çekme refleksinde olduğu gibi bir çift antagonistik kası) kontrol ettiği varsayılırdı. Öğrenilmiş davranışın örneğin sıçanların bir labirentte yol bulması böyle refleks devrelerinin zincirleme bağlanması sonucunda olduğu sanılırdı. Bu kabul edilebilir davranış organizasyonu modeli, bu yüzyılın büyük kısmında bilim adamlarında egemendi.
Böcek Uçuşu
Karmaşık davranışın refleksler zincirinden oluştuğu fikri, 1960’larda Stanford Üniversitesi’nden Donald Wilson’un araştırmasıyla çürütüldü. Wilson, çekirgenin uçmasını sağlayan devreler üzerinde çalışıyordu. Bu, refleks zincirleri olarak kabul edilebilecek iyi bir adaydı. Zira bir grup uçma kası, kanatları kaldıracak; bir süre sonra diğer bir gruptaki gerilme reseptörleri uyarılarak kanatları aşağı indirecek kasılmaları başlatacaktı. Kanatlar aşağı inince, yukarı kaldıran kas grubundaki gerilme reseptörleri, artan gerilme ile uyarılacaklar ve bir süre sonra “kaldırma refleksini harekete geçirecekti.” Bundan sonra kanatlar yukarı kaldırılacak ve bu defa aşağı indirici kas grubundaki reseptörler uyarılıp “indirme refleksini” başlatacaklar ve bu böylece sonsuza kadar devam edecekti.
Wilson, devreyi izleyerek gerekli gerilme reseptörlerini keşfetti ve reseptörlerden torasik gangliyonlara giden sinirleri kesti. Wilson, başlatıcı elemanların devreden uzaklaştırılmasıyla davranışın ortadan kalkacağını umuyordu. Fakat çekirge neredeyse normal uçuşunu sürdürerek onu şaşırttı. Yine de operasyonun bir etkisi olmuştu.
Kanat çırpma hızı hafifçe düşmüş ve koordinasyon bir dereceye kadar bozulmuştu. Fakat buna karşın davranış sürmekteydi. İzleyen araştırmalar, çekirgenin, gerilme reseptörleri olmaksızın da uçma yeteneğini  sürdürmesinden bir başka devrenin sorumlu olduğun ortaya koydu. Bu, torasik gangliyonlarda bulunan bir motor program ve gerilme reseptörlerinden gelen herhangi bir dış uyarıya gerek olmadan, uçmak için gerekli komutları üreten bir devreydi. Kuşkusuz normalde bu ayarlanmaktaydı.
sürdürmesinden bir başka devrenin sorumlu olduğun ortaya koydu. Bu, torasik gangliyonlarda bulunan bir motor program ve gerilme reseptörlerinden gelen herhangi bir dış uyarıya gerek olmadan, uçmak için gerekli komutları üreten bir devreydi. Kuşkusuz normalde bu ayarlanmaktaydı.
Wilson’un keşfinden bu yana omurgalılarda ve omurgasızlarda ayrıntılı bir biçimde incelenen birçok karmaşık, ritmik ya da sıralı davranışın, özelleşmiş bazı bağımsız devreler tarafından gerçekleştirildiği bulundu. Bu devreler, kas hareketlerini doğru sırada ve doğru zamanlamayla düzenlerler. Refleks zincirleri üzerine kurulu hiçbir karmaşık davranış bilinmemektedir. Motor programlar tarafından kontrol edilen davranışlar listesinde beslenme, yürüme, yüzme, kaşınma, ses çıkartma, yuva yapma, hücum etme, kur yapma ve çiftleşme davranışları yer alır.
Aplysia’da Beslenme
Araştırma teknikleri geliştikçe dikkatler, karmaşık davanışlardan sorumlu devrelerin hücre nörofizyolojik haritalanması üzerinde yoğunlaştı. Günümüzde iyi anlaşılmış motor program ağlarının çoğu yer değiştirme hareketinde gördüğümüz gibi ritmik ve tekrarlanan hareketleri içerir. Pratik gereksinimden dolayı araştırmalar, düzgün bir biçimde dakikalar hatta saatler boyu süren tipte davranışlar üzerinde odaklanmışır. Böylece bilimadamları, aynı devredeki iki hücreyi bulmak, bunlar arasındaki bağlantının doğasını tespit etmek, aktivitelerini aynı anda kaydetmek ve nihayet bunları işaretlemek (genellikle boya enjeksiyonuyla) için yeterli zamanı bulabilirler. Doğal olarak devrede ne kadar çok hücre varsa ve bunlar arasındaki ilişki ne kadar ayrıntılıysa, devrenin haritasını çıkartmak ve onu anlamak o kadar zaman alacaktır.
Neredeyse tamamen haritalanmış bir davranış yoluna güzel bir örnek Aplysia’daki beslenmeyi kontrol eden devredir. Bu devre, bilgi akışındaki dört adımı -fark etme, iletme, işleme ve cevap- nispeten karmaşık bir davranış bağlamında tanımlar. Yosunlar üzerinde beslenen Aplysia’nın kafasında, potansiyel besini tespit eden kemosensitif dokunaçlar bulunur. Bu reseptörlerden gelen impulslar analiz için Aplysia’nın beynine iletilirler. Eğer koku işleme ağı, hayvanın uygun bir besine temas ettiğine karar verirse (yumuşakçalar besinin kokusunu öğrenebilirler), beslenme davranışını kontrol eden internöronlara sinyaller gönderir. Fakat bu internöronlar, beynin diğer kısımlarından beslenmeyi önleyici sinyaller de alırlar. Örnegin kaçma, beslenmenin önüne geçer ve böylece hayvan, bir tehlike sezerse beslenme gibi düşük öncelikli aktiviteler durdurulur. Beslenme sürdürüleceği zaman bir motor program aktive edilir. Birçok omurgasız davranışında görüldüğü gibi merkezi sinir sistemi sadece bir gangliyondaki uygun bir devreyi inhibe etmeyi durdurur.
Beslenme 22 kas, altı motor nöron tarafından kontrol edilir. Bu motor nöronlar da protraktor ve retraktör olmak üzere “komut veren” internöronların iki antagonistik grubu tarafından koordine edilir. 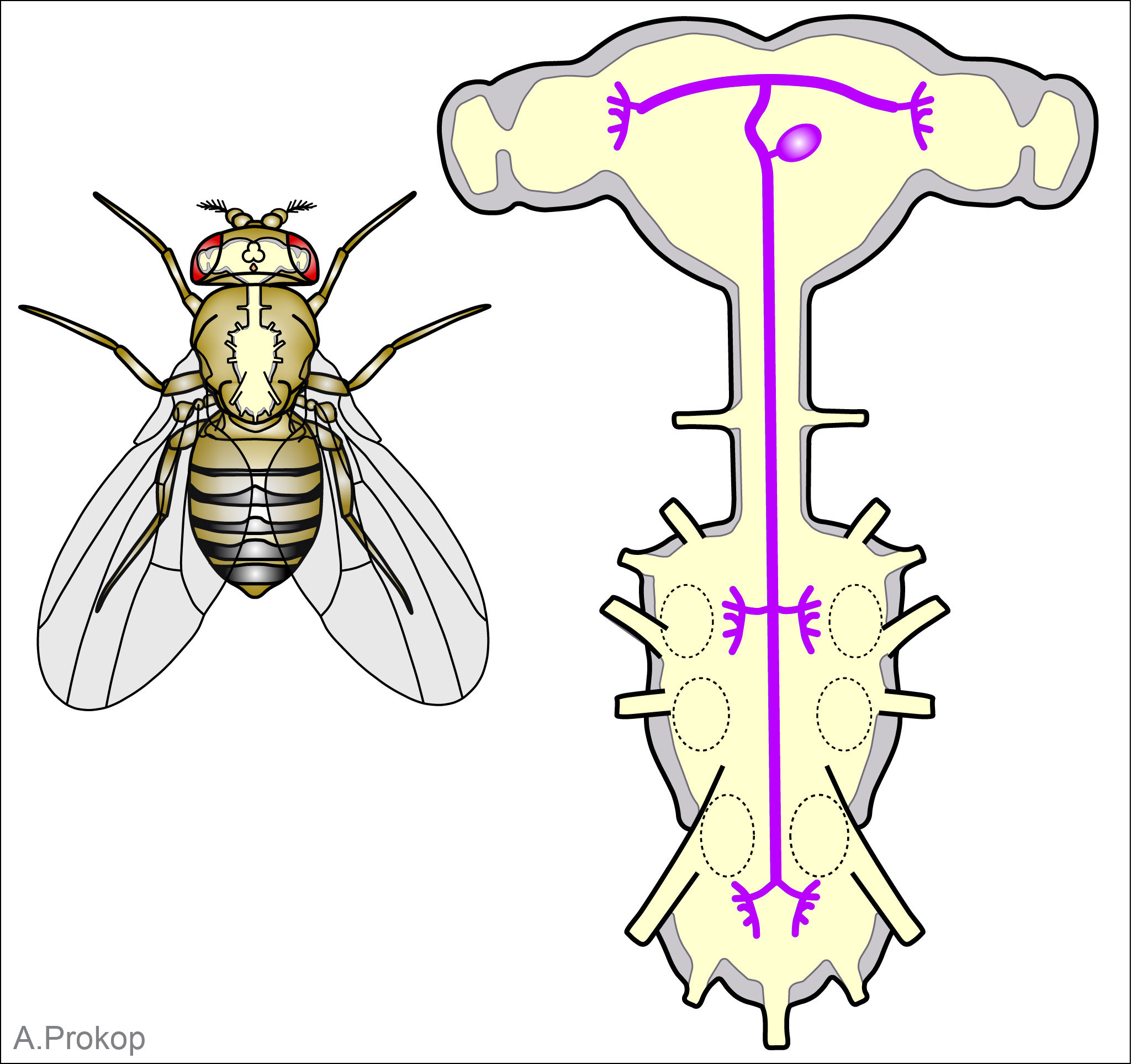 Protraktor ve retraktor kaslar, sırasıyla ekstensör ve fileksör kaslara analogtur. Protraktor internöronlar hey üç ya da dört saniyede kendiliklerinden ritmik impulslar çıkartırlar. Bu hücrelerden çıkan sinyaller ağzı açan ve “dişleri” besini ısıracak pozisyona getirmek için aşağı indiren kasları belli bir sırada aktive ederler. Bu hücreler aynı zamanda retraktor motor nöronları ve bunların komut internöronlarını inhibe ederler.
Protraktor ve retraktor kaslar, sırasıyla ekstensör ve fileksör kaslara analogtur. Protraktor internöronlar hey üç ya da dört saniyede kendiliklerinden ritmik impulslar çıkartırlar. Bu hücrelerden çıkan sinyaller ağzı açan ve “dişleri” besini ısıracak pozisyona getirmek için aşağı indiren kasları belli bir sırada aktive ederler. Bu hücreler aynı zamanda retraktor motor nöronları ve bunların komut internöronlarını inhibe ederler.
Protraktor internöronların ritmik döngüsü bunları sesiz faza getirince retraktor internöronları inhibe etmeyi durdururlar. İnhibisyon olmadığı zaman sürekli olarak impuls çıkartan bu hücreler, bu aşamada retraktor motor nöronları aktive edip protraktor kasları kontrol edenleri inhibe ederler. Böylece kasların kasılma sırasını, ısırmak ve ağızı kapatmak yönünde değiştirirler.
Alpysia’nın beslenme devresi, tipik bir motor programının üç temel özelliğini sergiler: Bunlar;
1.Program, çeşitli kasların hareketini koordine eden bağımsız bir devre tarafından çalıştırılır.
2.Bağımsız olmakla birlikte duyu geri bildiriminden faydalanarak davranışın otomatik olarak ince ayarını yapar. Bu örnekte uygun olduğunda ağzı açma fazını uzatıp güçlendirir.
3.Beynin doğrudan merkezi kontrolü altındadır. Aplysia’da beyin, kimyasal duyarlılığı olan hücrelerden ve “açlık” reseptörlerinden gelen bilgiyi entege eder ve gerektiğinde bu motor programı aktive eder. Benzer biçimde, diğer davranışlar gerekli olduğunda -örneğin bir avcı denizyıldızından kaçış- beyin beslenme davranışını durdurabilir. Beyin aynı zamanda, davranış hızını da değiştirebilir; gerektiğinde döngü süresini uzatabilir ya da kısaltabilir.
Aplysia’nın protraktorları ve retraktorları gibi iki grup komut hücresini kullanma stratejisine resiprokal inhibisyon adı verilir. Bu yöntem, görünürde tüm ritmik davranışların organize yolu olarak düşünülür. Motor programlar aynı zamanda yutma, gülme gibi ritmik olmayan davranışları da gerçekleştirir. Çok daha karmaşık davranışların düzenlenmesini sağlayan strateji, örneğin bir kuş yuvasının yapılması ya da bir kedinin bir fareye sessizce yaklaşması, onu kovalaması, yakalaması, öldürmesi parçalaması ve yemesi birçok ayrı motor programların kurulmasını ve bunları koordine edecek üst düzey devrelerin oluşmasını gerektirir. Bir organizmanın davranışlarının Alplysia’dakilere benzer, birbirleriyle etkileşen sinir devreleri grupları tarafından başarıldığının anlaşılması, modern nörofizyolojinin en büyük başarısıdır.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç