
Bundan yüzyılı aşkın bir süre önce, sinir impulsunun elektriksel değişiklikler içerdiği keşfedilince bilim adamları impulsun, tıpkı tellerden akan diğer akımlar gibi bir nöron boyunca iletilen basit bir elektrik akımı olduğunu düşündüler. Daha sonra açığa çıktı ki bir sinir impulsunun hızı, bir teldeki elektriğinkinden çok daha yavaştı. Bunun da ötesinde aksonun sitoplazmik özünün, basit elektrik akımlarına karşı direnci o kadar büyüktü ve akson zarı yeteri kadar iyonun dışarı kaçmasına izin veriyordu, ki pasif sinir impulsunun sadece birkaç milimetre yol aldıktan sonra sona ermesi beklenirdi. Buna karşı eğer bir nöronun aksonu boyunca çeşitli noktalarda sinir impulsunu ölçersek aynı kaldığını, yani gücünün yol boyunca azalmadığını buluruz. Diğer yandan, bir aksonu ezmek ya da zehirlemek, elektriksel iletim özelliğini değiştirmese bile impuls iletim yeteneğini yok edebilir. Kısaca, impuls iletimi canlı hücredeki aktiviteye bağımlıdır. Bu yüzden impuls, basit bir elektrik akımı olmaktan çok, nöron boyunca yayılan bir elektrokimyasal değişikliktir.
Sinir aktivitesinin modern teorisinin temel sınırları, 1902’de Almanya’daki Halle Üniversitesi’nden Julius Bernstein tarafından önerildi. Kendisi, bir sinir hücresinin içindeki ve onu çevreleyen sıvılardaki bazı iyonların derişiminin çok farklı olduğunu biliyordu. Sodyumun (Na+) derişimi hücre içinde çok düşükken, potasyumun (K+) derişimi çok yüksekti. Bu iyonik farklılık, daha düşük düzeyde olmak üzere hemen bütün hücrelerde vardır. Beinstein, aynı zamanda, negatif iyonlar için de (artık bunların hücre içindeki proteinler ve hücre dışı sıvıdaki klor iyonları olduğu biliniyor) aynı derecede önemli bir rol önerdi. Sonradan sinir hücresi zarının bu maddeler için neredeyse hiç geçirgen olmadığı ve Na+ ve K+ hareketinin sinir impulsu üretmede baskın rol aldıkları anlaşıldı. Bernstein, iyonların eşit olmayan dağılımının zarın iki yüzeyi arasında elektrostatik bir gradiyentle sonuçlandığını fark etti. Bunun sonucunda hücrenin iç tarafı dışarıya göre -70 mv’lık bir yüke sahip olur. Her ne kadar o günlerde zarın işlevi tam 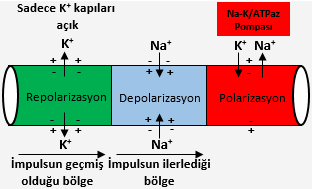 olarak anlaşılmaktan uzaktaysa da, Bernstein, nöron zarının farklı iyonlara karşı geçirgenliğinin farklı olduğunu ve iyonlarırı birbirinden ayrılmasının ve sonuçta oluşan elektrostatik gradiyentin nedeninin zarın bu büyük seçiciliği olduğunu önerdi. Bernstein, ayrıca bir impulsun geçişi sırasında zarın seçiciliğinin geçici olarak bozulduğunu da ileri sürdü. Dolayısıyla iyonlar serbestçe hareket edebilmekte ve zarın iki yüzeyi arasındaki elektrostatik gradiyent sıfıra düşmekteydi. Diğer bir ifadeyle zar, geçici olarak yükleri ayırma özelliğini kaybetmekteydi.
olarak anlaşılmaktan uzaktaysa da, Bernstein, nöron zarının farklı iyonlara karşı geçirgenliğinin farklı olduğunu ve iyonlarırı birbirinden ayrılmasının ve sonuçta oluşan elektrostatik gradiyentin nedeninin zarın bu büyük seçiciliği olduğunu önerdi. Bernstein, ayrıca bir impulsun geçişi sırasında zarın seçiciliğinin geçici olarak bozulduğunu da ileri sürdü. Dolayısıyla iyonlar serbestçe hareket edebilmekte ve zarın iki yüzeyi arasındaki elektrostatik gradiyent sıfıra düşmekteydi. Diğer bir ifadeyle zar, geçici olarak yükleri ayırma özelliğini kaybetmekteydi.
Buna ek olarak bu depolarizasyon, bazı yollarla komşu bölgelerin depolarizasyonunu da tetiklemekteydi. Bernstein’e göre sinir impulsu zar boyunca hareket eden bir depolarizasyon dalgasıydı. Bernstein’in hipotezi büyük ölçüde kabul gördüyse de uzun yıllar boyunca ne bu hipotezi destekleyen ne de bunun aleyhine hiç deneysel veri elde edilmedi. Sinir telleri, impuls iletimi sırasında içlerinde meydana gelen değişiklikleri tam olarak ölçebilmek için çok küçüktü.
1933’te Oxford’tan J. Z. Young, mürekkepbalığının çapı 1 milimetreye kadar varan çeşitli dev sinir telleri olduğunu keşfetti. Bu teller hayvanın kaçma tepkisine aracılık ederler. Bunlar, mürekkepbalığının gövde duvarı boyunca uzanıp hayvanın kendisini büyük bir hızla geriye doğru itmesini sağlayan kasları sinirlendirirler. Kaslar, suyu başa yakın bir huni aracılığıyla manto boşluğundan şiddetle dışarı atarak başarırlar. Mürekkepbalığının sinir tellerinin bu kadar kalın olması, sinir impulsunun çok hızlı iletilmesini sağlar. Genel olarak sinir telinin çapı, ne kadar büyükse iletim hızı da o kadar yüksektir. Mürekkepbalığındakiler kadar olmasa bile normalden kalın tellere, diğer birçok omurgasızda, çok hızlı iletimin önemli olduğu sinir devrelerinde rastlanır. Mürekkepbalığındaki teller özellikle çok kalındır. Çünkü mesajların katetmek zorunda olduğu uzaklık, omurgasızlarının çoğundakinden oldukça uzundur. (Omurgalılarda iletim hızını arttırmada alternatif bir adaptasyon olarak  miyelinli teller evrimleşmiştir.) Mürekkepbalığının dev sinir tellerinin keşfi, en sonunda biyologların iletim sırasında meydana gelen olaylaları ayrıntılı olarak çalışmalarına olanak sağlamış ve nörofizyolojide yeni bir devir açmıştır.
miyelinli teller evrimleşmiştir.) Mürekkepbalığının dev sinir tellerinin keşfi, en sonunda biyologların iletim sırasında meydana gelen olaylaları ayrıntılı olarak çalışmalarına olanak sağlamış ve nörofizyolojide yeni bir devir açmıştır.
1939’da Woods Hole, Massachussetts’den Flj. Curtis ve K. S. Cole ile Plymouth, İngiltere’den A. L. Hodgkin ve A. F. Huxley, mürekkepbalığının dev sinir teline çok ince bir elektrodu yerleştirmek üzere bir teknik geliştirdiler. Tuz çözeltisi ya da metalle dolu bir cam borudan oluşan bu mikroelektrot telin bir ucundan sokulup 1-3 cm ilerletildi. Elektrot bu şekilde yerleştirilmişken bir sinir impulsunun geçişi sırasında zarın iki yüzeyi arasındaki elektrostatik gradiyentin değiştiğini, Bernstein’in tahmin ettiği gibi göstermek mümkün oldu. Fakat doğru olmayan bir şey vardı: Gradiyent çok fazla değişmişti. Bir an için telin iç kısmı gerçekten dışarıya göre pozitif oldu. Daha sonra ise zar depolarize olmayıp bir an için polarizasyonunu tersine döndürdü. Uyarı, Bernstein’in daha önce düşünmüş olduğu gibi sadece zarın seçiciliğini ortadan kaldırmakla kalmadı; zira eğer buna yapsaydı zar potansiyeli kolayca sıfıra ulaşırdı. Bunun yerine, bu seçiciliği radikal bir biçimde değiştirdi. Hodgkin ve Bernard Katz, 1947’de zarın önce dışarı çıkmalarına izin verdiği iyonlardan çok daha fazla Na+ iyonunun hücreye girmesine izin vermesi gerektiğini önerdiler. Şimdi artık klasik olan uzun deney serilerinde Hodgkin ve Huxley, mürekkepbalığında impuls iletimi sırasında meydana gelen iyonik iletim değişikliklerinin ayrı ntılı kantitatif tanımları üzerinde çalıştılar. Onların bulguları tüm sinir hücreleri için geçerli bulundu. Hodgkin-Huxley modeline göre, dinleme durumundaki nöronun zarı polarizedir ve içi dışarıya göre negatiftir. Na+ iyonlarının derişimi dışarıda çok daha fazla, K+ iyonunu derişimi ise içerde daha fazladır. Bu durumda zarı bir elektrotla yapay olarak depolarize etmek zarın Na+ iyonlarına olan geçirgenliğinde büyük; fakat kısa süreli bir artışa yol açar. Bu iyonlar da hem yüksek derişimden düşük derişime doğru hareket etmelerini sağlayan doğal eğilimleri, hem de hücre içindeki net negatif yüklerin çekimi nedeniyle, zarı geçip hücre içine hücum ederler. Na+’nın içeri akışı o kadar çoktur ki bir an içerisi, gerçekten dışarıya göre pozitif olur. (Bu olur çünkü zardaki elektrostatik gradiyent sıfır hatta biraz pozitif olsa bile Na+’nın ozmotik derişimi içeriye göre daha yüksek kalmaya devam eder). Saniyenin kesiri kadar bir süre sonra, zarın Na+ geçirgenliği normale dönerken, K+’ya geçirgenliği büyük oranda artar. Şimdi K+ iyonları hücre dışına hücum ederler; çünkü bunların içteki derişimi dıştakinden büyüktür ve hücre içinde bir anda yükselen pozitif yük tarafından itilirler. Pozitif yüklü iyonlarının dışarı çıkışı, hücre içindeki yükü başlangıçtaki negatif değerine geri döndürür. Kısaca, zarın iç yüzeyi başlangıçta negatiftir, Na+ iyonları içeri hücum edince pozitif olur ve sonra K+ iyonları dışarı akın edince yeniden negatif hale gelir.
İmpuls nöron boyunca iletilir, çünkü her noktada meydana gelen bu değişiklikler döngüsü zarda hemen yanındaki noktayı depolarize eder ve burada benzer geçirgenlik değişikliği döngüsünü başlatır ve bu da yanındaki noktada yeni döngüyü başlatarak devrilen dominolar zinciri gibi aksonda ilerlemesini sağlar. Hodgkin-Huxley impuls iletim modeli sinir impulsunun neden, bir telde ilerleyen basit bir elektrik akımının aksine sinir teli boyunca ilerlediği sırada gücünde bir azalma meydana gelmediğini açıklar. İmpuls kendini yeniler, birbirini izleyen her noktadaki elektriksel hücre zarı değişiklik, ya da aksiyon potansiyeli yeni bir olaydır ve bunun büyüklüğü sitoplazma  önceki noktalarda meydana gelen elektriksel olaylarla aynıdır. Bir pil ya da dinamo tarafından üretilen bir elektrik akımı, bir tel tarafından pasif olarak taşınır; fakat bir sinir impulsu, enerjisini üzerinde ilerlediği yoldan alır ve tel üzerinde birbirini izleyen her noktada bunu yeniler. (Miyelinli teller impulsu aynı kalınlıktaki miyelinsiz tellere göre daha hızlı iletirler; çünkü aksiyon potansiyelinin oluştuğu noktalar zarın birbirine komşu bölgeleri değil, birbirini izleyen düğümlerdir. Miyelin, iyonların giriş çıkışını önlemek yoluyla, bir düğümde meydana gelen zar potansiyelindeki değişikliğin bir sonraki düğüme sıçramasını sağlar.)
önceki noktalarda meydana gelen elektriksel olaylarla aynıdır. Bir pil ya da dinamo tarafından üretilen bir elektrik akımı, bir tel tarafından pasif olarak taşınır; fakat bir sinir impulsu, enerjisini üzerinde ilerlediği yoldan alır ve tel üzerinde birbirini izleyen her noktada bunu yeniler. (Miyelinli teller impulsu aynı kalınlıktaki miyelinsiz tellere göre daha hızlı iletirler; çünkü aksiyon potansiyelinin oluştuğu noktalar zarın birbirine komşu bölgeleri değil, birbirini izleyen düğümlerdir. Miyelin, iyonların giriş çıkışını önlemek yoluyla, bir düğümde meydana gelen zar potansiyelindeki değişikliğin bir sonraki düğüme sıçramasını sağlar.)
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç