

Terminal sisterna – Sarkoplazmik retikulumun Z diskine yakın olan parçasıdır. Triad = T tubule + terminal sisterna.
ATP’nin kaslarda miyozin filamentlerinin çark hareketine yakıt oluşturduğunu biliyoruz. Kasların bu yakıtı ne zaman “yakacaklarını” nasıl bildiklerini ve nasıl olup da sürekli olarak kasılı kalmadıklarını merak edebilirsiniz. Sinir ve endokrin sistemlerinde görmüş olduğumuz gibi, kimyasal kontrol çeşitli düzeylerde yapılabilir. Kaslarda kritik adım, miyozin başlarının aktin filamentlerine bağlanmasıdır. Öyleyse nasıl oluyor da bir sinir uyarısı bir sinir—kas kavşağında (bağlantı bölgesinde) miyozin başlarının kasılmayla sonuçlanan moleküler bağlanmayı başlatıyor ve kontrol ediyor?
Dinlenme durumundaki bir nöronun zarı gibi, dinlenme durumundaki bir kas telinin zarı da polarizedir; dış yüzey iç yüzeye göre artı yüklüdür. Sinir-kas kavşağında bir sinir aksonundan salınan uyarıcı transmitter madde kas zarında bir anlık depolarizasyona yol açar. Eğer depolarizasyon eşik değer düzeyine ulaşırsa, aynen sinir hücrelerinde görülen voltaj-kapılı Na+ ve K+ kanallarının kombinasyonuyla bir impuls ya da aksiyon potansiyeli oluşur ve telin yüzeyi boyunca iletilir. Aksiyon potansiyeli kasılma işlemini dolaylı olarak, Ca++ iyonları aracılığıyla aktive eder.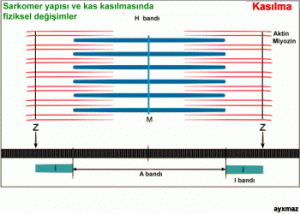
Kaslarda kasılmanın iyonik kontrolüyle ilgili ilk ipucu, 1949’da Pennsylvania Üniversitesi’nden L. V. Heilbrunn ve Floydi Wiercinski kas tellerine çok çeşitli maddeler enjekte edip bunlardan sadece bir kalsiyum tuzunun kasılmaya yol açtığını buldukları zaman elde edildi. Dolayısıyla, kalsiyum iyonlarının (Ca++) (ki normalde bir kısmı uyarılan kas fibrelerinin içine akar) çapraz köprüleri aktive ettiğini söylemek mantıksız olmaz. Buna karşın, bu fikir ilk defa ileri sürüldüğünde buna karşı iki büyük itiraz geldi. Bunlardan birincisi omurgalı kasının kasılmasının çok sayıdaki miyofibrilin aynı anda kısalmasını gerektirdiği; fakat telin ortasındaki miyofibrillerin yüzeyden uzak oldukları ve dolayısıyla dışarıdan gelen Ca++ iyonlarının, telin uyarılmasıyla kasılması arasındaki o kısa süre içinde yeteri kadar hızlı difüze olamayacaklarıydı. İkinci olarak da hücreye, hızlı ve çok sayıda sinir uyarıları sonucunda meydana gelen sürekli kasılmalardan sorumlu olabilecek kadar çok Ca++ girmediğini gösteren deliller vardı .
1955’de kas tellerinde yoğun bir tübül ağının bulunduğu yeniden keşfedilince, uyarımın kasılmaya bağlanması problemine de çözüm yolu açıldı. ‘Yeniden keşfedilme’ diyoruz; çünkü tübüller Birinci Dünya Savaşından önce histologlar tarafından biliniyordu; fakat Rockefeller Enstitüsünde çalışan Stanley Bennett ve Keith R. Porter, o zamanlar yeni bir teknik olan elektron mikroskopla bunları yeniden tespit edinceye kadar unutulmuşlardı. Kısa sürede, tübüllerin iki ayrı; fakat işlevsel olarak ilişkili sistem içerdiği bulundu: dışarıya açılmayan sarkoplazmik retikulum ve teli sarmalayan plazma zarının bir kısmı olan T sistemi ya da transvers tübül sistemi.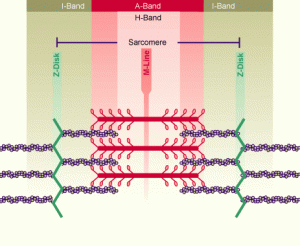
Sarkoplazmik retikulum, tüm hücrelerde bulunan endoplazmik retikulumun kas hücresindeki büyük ölçüde özelleşmiş bir tipidir. Zardan oluşan kanalları, miyofibrilde sarkomerin çevresini saran bir ağ oluştururlar. Bir sarkomerin uzak ucundaki sarkoplazmik retikulum ile bundan sonra gelen sarkomerin yakın ucundaki birbirlerine çok yakın yer alırlar; fakat bunların arasında, Z çizgisi hizasında, T sisteminin bir tübülü (memelilerde iki tübül) uzanır.
Her ne kadar sarkoplazmik retikulum ve T tübülleri doğrudan temastaysalar da, bunların lümenleri (iç boşlukları ) arasında bağlantı yoktur ve böylece bunların içerikleri karışmaz. Kas hücresinin yüzeyinde bir aksiyon potansiyeli iletildiği zaman, bu, T tübüllerinin zarları yoluyla telin içine de girer. Aksiyon potansiyeli, difüze olan iyonlardan çok daha hızlı hareket eder. Bu hızlı hareket sayesinde kasılma için verilen uyarı tüm miyofibrillere hemen hemen aynı anda ulaşır ve böylece yüzeye yakın miyofibrillerle telin merkezindekiler birlikte kasılabilirler.
T tübülleriyle sarkoplazmik retikulum arasındaki bu yakın bağlantı, T tübulün zarı boyunca ilerlemekte olan aksiyon potansiyellerinin retikulumu tetiklemesini sağlar. Retikulum büyük miktarlarda Ca++ iyonu içerir ve bunlar da, Heilbrunn ve Wiercinski’nin buldukları gibi, miyofibrillerin kasılmasını sağlarlar. Aksiyon potansiyeli, retiküler zarların Ca++ geçirgenliğinde ani ve belirgin bir artışa yol açarak büyük miktarda iyonun retikulumu terketmesini sağlar. Kasılmayı doğrudan uyaran, bu aniden serbestleşen hücre içi Ca++’dur.
Şimdi, soru şudur: Kalsiyum, kas tellerinin kasılmasını nasıl tetikler?
Cevap vermek için, ince filamentlerin yapısına daha yakından bakmak zorundayız. Daha önce gördüğümüz gibi, ince filamentin esas proteini aktindir. Buna ek olarak, bu filamentler önemli düzenleyici proteinler olan tropomiyozin ve troponin kompleksi içerirler.
Aktin molekülünün alt birimleri globülerdir ve birbirine heliks biçiminde dolanmış iki sıra oluştururlar. Bu sıralar boyunca da ilk düzenleyici protein olan tropomiyozinin ince uzun molekülleri uzanır. Dinlenme durumundaki kasta, tropomiyozin, kalın filamentteki çapraz köprülerin aktine bağlanmasını önler (büyük bir olasılıkla bunun üzerindeki miyozinin bağlanma bölgelerini örterek).
Diğer düzenleyici proteinler olan troponin kompleksinin molekülleri de globüler olup, her yedinci aktin biriminin yakınında, üçlü birimler halinde yer alırlar. Her kompleksin üç bağlama bölgesi vardır: biri aktin, biri tropomiyozin ve biri de Ca++ iyonları için. Ca++ iyonları sarkoplazmik retikulumdan salınınca, troponin kompleksinin kalsiyum bağlayıcı bölgelerine bağlanırlar ve troponin bu bağlanmaya yapısal bir değişiklikle tepki gösterir. Bu değişiklik, tropomiyozinin pozisyonunda bir kaymaya yol açar ve bunun sonucunda da tropomiyozin aktini inhibe etmeyi durdurur. Böylece aktin, miyozinin çapraz köprülerinin bağlanması için serbest hale gelir ve kasılma işlemi başlatılır. Şimdi artık, çapraz köprü hareketini ve bunun uyarılmasını içeren geçerli modeli anlamak için tüm elemanlara sahibiz. Dinlenme durumundaki bir kasta, çapraz köprüler-kalın filamentlerin globüler miyozin başları- ATP tarafından aktive edilmiş (dik) durumdadırlar; fakat ince aktin filamentlerine bağlanamazlar; çünkü tropomiyozin aktin moleküllerinin üzerindeki bağlanma bölgelerini inhibe etmektedir. Bir motor sinirden gelen uyarım bir aksiyon potansiyeli oluşturup bu da T tübüller boyunca iletilip kas tellerinin iç kısmına ulaşınca, sarkoplazmik retikulum Ca++ iyonları salar. Bu iyonların bir kısmı troponin kompleksine bağlanır ve bunun bir yapı değişikliği geçirmesine yol açar. Böylece tropomiyozin bulunduğu yerden ayrılır ve aktin üzerindeki miyozin bağlanma bölgeleri, çapraz köprülerin bağlanması için açığa çıkar. Miyozinin bağlanması çapraz köprülerin bükülmesine yol açar ve böylece, filamentlerin birbiri üzerinde kaymasını sağlayan darbe gücü başlatılır. Aktinle bağlantısı koptuktan sonra bir miyozin çapraz köprüsüne yeni bir ATP molekülü bağlanınca, baş, yeniden ilk dik durumuna getirilir.
Bu iyonların bir kısmı troponin kompleksine bağlanır ve bunun bir yapı değişikliği geçirmesine yol açar. Böylece tropomiyozin bulunduğu yerden ayrılır ve aktin üzerindeki miyozin bağlanma bölgeleri, çapraz köprülerin bağlanması için açığa çıkar. Miyozinin bağlanması çapraz köprülerin bükülmesine yol açar ve böylece, filamentlerin birbiri üzerinde kaymasını sağlayan darbe gücü başlatılır. Aktinle bağlantısı koptuktan sonra bir miyozin çapraz köprüsüne yeni bir ATP molekülü bağlanınca, baş, yeniden ilk dik durumuna getirilir.
Sinirin kası uyarmaya devam ettiği süre içinde, serbest Ca++ iyonları (ve ATP) mevcut olduğu sürece çapraz köprü bağlanması, darbe gücü ve eski haline geri dönme döngüsü, kas kasılmayı sürdürdükçe, tekrar tekrar meydana gelir. Fakat eğer sinirsel uyarım durursa kas gevşer; çünkü sarkoplazmik retikulumun zarında bulunan kalsiyum pompası , Ca++’u hı zla retikuluma geri alır ve troponin-tropomiyozin sistemi aktin üzerindeki miyozinin bağlanma bölgelerini yeniden inhibe eder. Yeniden Ca++ salınması olup, yeni bir kasılmayı başlatacak aşamalar gerçekleşmeden, kas telinde artık ATP “yakılmaz”. Sonuçta, kasılmayı sürdürmek için bir bedel ödemeye gerek kalmaz- rigor mortis adı verilen olay bu noktayı iyi açıklar: Ölümden sonra sinir uyarım durunca kaslar önce gevşerler; fakat birkaç saat sonra, sarkoplazmik retikulum parçalanıp kalsiyumu serbest bırakınca tekrar kasılırlar ve vücudun katılaşmasına yol açarlar.
Bu tanım sadece omurgalı iskelet kası için geçerlidir. Birçok yönden benzerlik göstermesine karşın düz kas, bazı ilginç farklılıklar sergiler. İskelet kasında, ATP, miyozin başlarını aktive eder ve Ca++ iyonları, aktin filamentlerinin troponin kompleksine bağlanarak, hareketi başlatırlar. Düz kasta, ATP işe karışmadan önce, Ca++ iyonları miyozini iki aracı enzim yoluyla aktive ederler. Bu, düz kasın neden böyle yavaş hareket ettiğini açıklamada yardımcı olur. Aynı zamanda, düz kasların hormonlar tarafından aktive edilme kapasitesinden de sorumludur.
Hormonlar sıklıkla Ca++ iyonlarına özgü zar kanallarını açarak etki ederler. Bu iyonlar da, ikinci haberci rolüne uygun olarak, sitozolde bir enzim kompleksine, örneğin kalinodülin, bağlanıp onu aktive ederler. Düz kasta meydana gelen olay tam olarak budur. Burada kalmodülin, troponinin iskelet kasındaki rolünü üstlenir. Gerçekten de, troponin kompleksinin üç alt biriminden biri modifiye olmuş bir kalmodülin molekülüdür.
Endokrin sistemle omurgalı düz kasları arasında bir ilişki daha vardır: siklik AMP miyozinin aktivasyonunda ikinci haberci rolü oynar. Düz kasın yararlı; ancak oransal olarak daha yavaş kasılma sisteminden iskelet kasının yüksek düzeyde organize olmuş hızlı kaşımasına geçişteki kritik evrimsel aşamalar şunlar olmalıdır:
(1) Kasılma birimlerini ayrı sarkomerlere bölen Z çizgilerinin ortaya çıkması,
(2) Kalmodilinin küçük bir modifikasyonla troponinin üç alt biriminden biri haline gelmesi,
(3) Endoplazmik retikulumun, miyofibrillere Ca++ taşınması için özelleşmiş bir sarkoplazmik retikuluma evrimleşmesi.
Kaynakça:
https://www.sciencedirect.com
Yazar: Taner Tunç